maestrowindu escribió:cada uno tiene lo que se merece es mi punto de vista
 Pues va a ser que no. Qué motivos tendría un supuestio Dios para dejar inválido a un niño de 5 años? Por decir algo...
Pues va a ser que no. Qué motivos tendría un supuestio Dios para dejar inválido a un niño de 5 años? Por decir algo...

maestrowindu escribió:cada uno tiene lo que se merece es mi punto de vista
kinglor escribió:Dios no existe, o si existe nuestro planeta lo creo de forma ilegal y por eso no controla nada. Espero que no venga un moderador divino y cierre nuesto hilo, huy! que diga nuestro planeta.
Na en serio, creo que lo único que puede explicar estas últimas y constantes catastrofes naturales es que nos estamos cargando el planeta y el pobre esta agonizando, defendiendose como todo organismo natural de los parasitos que lo estan infectando, asi que en cierta medida todos somos culpable de estas catastrofes, cuando digo todos me refiero a la humanidad como entidad comun, sin dividir en razas, paises o momentos historicos...
ernen escribió:Dios no existe y si existiera habria que buscarlo para rebentarlo de una paliza. xD
Es sencillo solo es un invento de marketing de una empresa llamada iglesia. Nada mas.
falkiño escribió:La fotosíntesis es un proceso que sintetiza glucosa a partir de dióxido de carbono y agua con la ayuda de la luz solar; en el proceso se libera oxígeno molecular como subproducto. Éso es fotosíntesis, lo demás e suna flipada tuya que no viene a cuento xD
"La Vitamina D es una prohormona, por lo que no tiene actividad hormonal por si misma, pero si se convierte a hormona activa 1,25-D a través de un mecanismo de síntesis muy regulado. La producción de vitamina D en la naturaleza, al parecer requiere de la presencia de rayos UV, incluso la vitamina D en alimentos comestibles es derivada en última instancia de organismos; desde setas u hongos a animales, los cuales no pueden sintetizarla sino a través de la acción de los rayos solares, en algún punto de la cadena sintética. Por ejemplo; los peces contienen vitamina D solo porque consumen algas del océano que sintetizan la vitamina D en aguas bajas por la acción de rayos UV.
La deficiencia de vitamina D causa también:
Osteoporosis: caracterizada por huesos frágiles.
Osteomalacia: una enfermedad que debilita los huesos en adultos y que es similar al raquitismo.[1]
Cuando el cuerpo carece de vitamina D es incapaz de regular adecuadamente los niveles de calcio y fosfato. Si los niveles sanguíneos de esos minerales disminuyen, las otras hormonas corporales pueden estimular la liberación de calcio y fosfato desde los huesos al torrente sanguíneo para elevar los niveles.
El raquitismo es una enfermedad ósea que afecta a personas cuando se presentan estas deficiencias que causan debilitamiento y reblandecimiento progresivo de la estructura ósea."[2]
Falkiño escribió:Yo no tomo CO2, H2O y luz solar para realizar la fotosíntesis y generar mis sustancias orgánicas; e igual que yo, ningún ser humano ni ser vivo del reino Animalia (o Metazoa) puede hacerlo
A nivel de oxidación en nuestro cuerpo también se producen procesos de fotosíntesis, pero me refiero especialmente a la luz del sol
...
En la fotosíntesis se libera oxígeno, que será utilizado en la respiración aerobia como oxidante...
Algunas sustancias al absorber luz, sufren cambios químicos; utilizan la energía que la luz les transfiere para alcanzar los niveles energéticos necesarios para reaccionar, para obtener una conformación estructural más adecuada para llevar a cabo una reacción o para romper algún enlace de su estructura (fotólisis).
La fotosíntesis en las plantas, que generan azúcares a partir de dióxido de carbono, agua y luz; la síntesis de vitamina D en la piel; la ruptura de dihalógenos con luz en las reacciones radicalarias o el proceso de visión en el ojo, producido por la isomerización del retinol con la luz, son ejemplos de reacciones fotoquímicas. El área de la química encargada del estudio de estos fenómenos es la fotoquímica.[3]
La fotólisis es la ruptura de enlaces químicos por causa de energía radiante. Se llama fotólisis o fotolisis, fotodisociación, o fotodescomposición a la disociación de moléculas orgánicas complejas por efecto de la luz, y se define como la interacción de una o más fotones con una molécula objetivo. Es el proceso en el que se basa la fotosíntesis.[4]
Prácticamente toda la vida sobre la Tierra depende de la energía solar, ya sean humanos (que dependen de los organismos fotosintéticos para obtener alimento) o plantas o algas fotosintéticas. Incluso en los humeros negros, en las profundidades abisales más oscuras, la vida depende del Sol. Los microorganismos que propician el crecimiento de los gusanos tubícolas gigantes necesitan sulfuros y oxígeno, la fuente del cual son, en última instancia, los organismos fotosintéticos que medran mucho más arriba. Por el contrario, todo lo que requieren los metanógenos de la Ciudad Perdida para sobrevivir es dióxido de carbono, agua líquida y peridotita, que reacciona para producir las materias primas necesariaas.[5]
"El Dr. Garza-Valdés afirma haber encontrado en muestras de la Sábana una capa de materia orgánica producida por microorganismos que todavía viven y se multiplican en los hilos. Según sus experimentos, esta especie de plástico adherido a las fibras es resistente a todos los métodos normales de limpieza, y puede constituir un porcentaje importante de materia carbonácea. Lo mismo ha sido afirmado por el Dr. Kouznetsov. Sin una serie de muestras bien controladas, y de medidas cuantitativas, no es posible juzgar la importancia que este material tendría para alterar la datación del lienzo (2).. Pero es sorprendente que se afirme (en el artículo original de "Nature") el mismo contenido de C 14 en muestras que se sometieron a limpieza muy profunda y en otras que se midieron sin limpieza alguna, a pesar del obvio estado de suciedad de la tela tras siglos de manipulación y exposición a humo, agua, y otros contaminantes."
Falkiño escribió:Puntal, mezclar respiración celular con fotosíntesis es como mezclar la mecánica de un coche con las mareas.
Una de las formas de verificar la actividad biológica es una curiosa propiedad de los sistemas celulares, como la fotosíntesis que incorporan CO2 de diversas fuentes para organificarlo. Existen dos isótopos estables del carbono, C12 y C13, siendo sus abundancias relativas fijas en la atmósfera. Cuando se incorpora CO2 por un sistema biológico, este prefiere ligerísimamente el isótopo más ligero, enriqueciendo las rocas carbonatadas en el otro isótopo.
Zachary Adam de la Universidad de Washington en Seattle afirma que procesos mareales mayores que los actuales, producidos por una luna situada a una distancia mucho menor podrían haber concentrado partículas radiactivas de uranio y otros elementos radiactivos en la marea alta en las playas primordiales donde debieron haber sido los responsables de generar los componentes elementales de la vida. De acuerdo con los modelos de computación publicados en Astrobiology un depósito de tales materiales radiactivos podría haber mostrado la misma reacción nuclear autosostenida que se encuentra en el yacimiento de Uranio de Oklo, en Gabón. Esta arena radiactiva proporciona suficiente energía para generar moléculas orgánicas, como aminoácidos y azúcares a partir de acetonitrilo procedente del agua. La monazita radiactiva también libera fosfatos solubles en las regiones que se encuentran entre los granos de arena, haciéndolos biológicamente accesibles. Así pues los aminoácidos, azúcares y fosfatos solubles pueden ser producidos simultáneamente, de acuerdo con Adam. Los actínidos radiactivos, que entonces se encontraban en mayores concentraciones, pudieron haber formado parte de complejos órgano-metálicos. Estos complejos pudieron haber sido importantes como primeros catalizadores en los procesos de la vida.
John Parnell de la Universidad de Aberdeen sugiere que tales procesos formaron parte del «crisol de la vida» en los comienzos de cualquier planeta rocoso hasta que éste fuera lo suficientemente grande para generar un sistema de tectónica de placas que aportara minerales radiactivos a la superficie. Puesto que se cree que la Tierra en sus orígenes estaba formada por muchas «microplacas», se darían condiciones favorables para este tipo de procesos.
Homoquiralidad:
Algunos procesos de la evolución química deberían explicar el origen de la homoquiralidad, es decir, el hecho de que todos los componentes elementales de los seres vivos tienen la misma quiralidad, siendo los aminoácidos levógiros, los azúcares ribosa y desoxirribosa de los ácidos nucleicos son dextrógiros, así como los fosfoglicéridos quirales. Se pueden sintetizar moléculas quirales, pero en ausencia de una fuente de quiralidad o de un catalizador quiral se forman en una mezcla 50/50 de ambos enantiómeros, a la cual se le llama mezcla racémica. Clark sugirió que la homoquiralidad pudo comenzar en el espacio, puesto que los estudios sobre los aminoácidos del meteorito Murchison mostraron que la L-alanina era dos veces más frecuente que la forma D, y el ácido L-glutámico era 3 veces más prevalente que su contrapartida dextrógira. Se ha sugerido que la luz polarizada tuvo el poder de destruir uno de los enantiómeros dentro del disco protoplanetario. Noyes ha demostrado que la desintegración beta provocaba la destrucción de la D-leucina en una mezcla racémica y que la presencia de 14C, presente en grandes cantidades en las sustancias orgánicas del ambiente temprano de la tierra, podría haber sido la causa. Robert M. Hazen ha publicado informes de experimentos realizados en distintas superficies cristalinas quirales que actuaban como posibles lugares de concentración y ensamblaje de monómeros quirales en macromoléculas. Una vez estabilizado el sistema, la quiralidad podría haber sido seleccionada favorablemente por la evolución. Los trabajos con compuestos orgánicos encontrados en meteoritos tienden a sugerir que la quiralidad es una característica de la síntesis abiogénica, puesto que los aminoácidos actuales son levógiros, mientras que los azúcares son predominantemente dextrógiros.[6]
Metano
Un equipo de investigadores estadounidenses ha logrado sintetizar metano, el principal componente del gas natural, en el laboratorio simulando las condiciones de presión y temperatura que reinan en el interior de la Tierra y Marte. Aunque al profano el experimento puede parecerle una insignificancia, la realidad es bien otra.
El metano surgido del submundo virtual da pábulo a la idea de que este gas podría en teoría alimentar colonias de microorganismos en la Tierra, así como en el Planeta Rojo, lo que ha despertado el interés de los exobiólogos. Y el estudio deja entrever la posibilidad de que existan vastas reservas de combustibles fósiles a profundidades hasta ahora no exploradas, lo que ha llamado la atención de las petroleras.
El metano es un hidrocarburo que guarda una íntima relación con otros combustibles fósiles, como el petróleo y el carbón. Los tres provienen de la descomposición de restos animales y vegetales que murieron hace cientos de millones de años, y que pasaron a formar parte de las llamadas rocas sedimentarias. Al menos, ésta la hipótesis defendida por la mayoría de los científicos, que otorgan al petróleo y demás hidrocarburos una génesis biológica. Ahora bien, no todos los expertos comulgan con esta idea. Ya en los años noventa, el recientemente fallecido Thomas Gold, astrofísico de la Universidad de Cornell, lanzó la posibilidad de que el petróleo tuviera un origen inorgánico y que surgiera de hidrocarburos, como el metano, que quedaron atrapados en el interior de la Tierra cuando se formó hace 4.500 millones de años. Estos gases migrarían desde el manto terrestre, que posee unos 2.895 kilómetros de espesor, hasta la superficie a través de las grietas presentes en la corteza, que se extiende hasta unos 33 kilómetros de profundidad. Ahora bien, una parte de los gases quedaron atrapados debajo de la superficie terrestre y, de ésta, una proporción considerable sufrió una serie de transformaciones químicas que dieron origen al petróleo y al carbón. Los planteamientos de Gold eran plausibles, pero tenían un punto débil, ya que no explicaban la presencia de moléculas biológicas en el petróleo.
Para salir al paso de los detractores, el astrofísico propuso que la materia orgánica era la huella de una actividad biológica dejaba por microbios que se alimentaban de petróleo e hidrocarburos. La idea de microbios "zampapetróleo" desató la risa de muchos, hasta que en el año 2002 un equipo de científicos estadounidenses informara de la presencia de microorganismos a 200 metros de profundidad, debajo del suelo de Idaho. Estos seres del averno comen hidrógeno y eructan metano en un ambiente con una temperatura por encima del punto de ebullición y a una presión infernal.
Para sintetizar el mismo metano, Henry Scott y sus colegas de la Universidad de South Bend han mezclado agua, óxido de hierro y calcita, y han sometido el cóctel a una presión salvaje y a una temperatura de 1.500 ºC, similares a las que hay en el manto superior terrestre. Resultado: el gas surgió de una reacción sencilla. La lectura de este ensayo, que acaba de ser hecho público por la National Academy of Sciences estadounidense, sugiere que el metano puede permanecer estable en regiones profundas de nuestro planeta que, según los cálculos, alcanzarían los 100 km. Las compañías petrolíferas generalmente no perforan más allá de los 8 kilómetros, lo que significa que a mayor profundidad podrían existir unas reservas de combustible fósil gigantescas, si tenemos en cuenta las dimensiones del manto terrestre. Obviamante, el principal problema estaría en cómo explotarlas.
El metano sintetizado por el equipo de Scott no sólo posee implicaciones en la economía y ecología terrestre, sino que tiene un efecto inmediato en la ciencia de la exobiología, que se dedica a la búsqueda de vida extraterrestre. Esto es así porque la presencia de metano en la atmósfera de un planeta alberga la posibilidad de que sea producto de la actividad biológica. En el mes de marzo, la nave Mars Mars Express, en órbita en el planeta rojo desde el pasado mes de diciembre, detectó este gas en la atmósfera marciana. Algunos científicos no tardaron en declarar de que podría ser una señal de vida, sobre todo porque el metano se mantiene por un corto período de tiempo en la atmósfera de Marte. Esto significa que tiene que ser reemplazado de manera constante de alguna forma. Y los dos productores de metano son los volcanes y los microbios. Si fuera a través de la actividad volcánica, la lava depositada en la superficie produciría el gas, pero hasta ahora no se han localizado zonas volcánicas en el planeta rojo. La otra alternativa es a través de microbios, ya que en la Tierra hay bacterias que producen gas metano del hidrógeno y el dióxido de carbono, y no necesitan oxígeno para desarrollarse. Ahora bien, el experimento de Scott ofrece una tercera génesis del metano; esto es, un origen químico en las profundidades marcianas. Pero no hay mal que por bien no venga: este metáno de padres inorgánicos podría servir de fuente de alimento para colonias de microorganismos que prosperaran en el infierno marciano.[7]
Las bacterias quimiotróficas necesitan de la presencia de oxígeno y no necesitan energía procedente de la luz solar. La energía empleada para hacer posible sus reacciones de síntesis se obtiene mediante oxidación de moléculas inorgánicas tales como compuestos de hidrógeno, azufre y hierro, o a partir de la oxidación de hidrógeno gaseoso.
Un quimioautótrofo es un organismo que puede crecer en un medio estrictamente mineral en la oscuridad, obteniendo carbono a partir del CO2, y su ATP y poder reductor de la respiración de un sustrato inorgánico.
La especificidad del sustrato de los quimioautótrofos permite establecer cinco subgrupos principales.
• Las bacterias nitrificantes utilizan compuestos reducidos de nitrógeno inorgánico como fuente de energía.
• Las bacterias oxidadoras de azufre emplean H2S, azufre elemental o sus óxidos parcialmente reducidos como fuente de energía.
• Las bacterias del hierro oxidan hierro y manganeso reducidos, pero no compuestos reducidos de azufre.
• Las bacterias del hidrógeno utilizan hidrógeno molecular como fuente de energía.
• Las carboxidobacterias utilizan monóxido de carbono.
http://www.drpez.net/panel/showthread.php?t=263632
¿Nuestro antepasado más antiguo fue una roca alimentada por protones?
Peter Mitchell fue un personaje excéntrico. Durante gran parte de su carrera trabajó en su propio laboratorio en una casa solariega restaurada en Cornwall, en el Reino Unido, y su investigación fue financiada en parte por su rebaño de vacas lecheras. Sus ideas sobre el proceso más básico de la vida —cómo se obtiene la energía— le parecían ridículas a sus colegas biólogos
Recuerdo que pensé que yo apostaría a que [la obtención de la energía] no funciona de esa manera”, escribió el bioquímico Leslie Orgel sobre su reunión con Mitchell, hace medio siglo. “Desde Darwin y Wallace la biología no ha tenido una idea tan no intuitiva como las de, por ejemplo, Einstein, Heisenberg y Schrödinger”.
En las décadas siguientes, sin embargo, quedó claro que Mitchell tenía razón. Su reivindicación fue completa cuando ganó un premio Nobel en 1978. Incluso hoy en día, sin embargo, la mayor parte de los biólogos no han comprendido aún las implicaciones de sus revolucionarias ideas, especialmente sobre el origen de la vida.
“Las ideas de Mitchell eran sobre cómo se organizan las células en el espacio, y la generación de energía celular es una función de eso”, dice Mike Russell, geoquímico del Jet Propulsion Laboratory de la NASA en Pasadena, California. “El problema es que la mayoría de las ideas sobre el origen de la vida no tienen en cuenta la organización espacial y el suministro de energía que maneje la reproducción o el crecimiento.”
Unos pocos investigadores, entre ellos: Russell, han estado replanteando el origen de la vida a la luz de las ideas de Mitchell. Creen que la mayoría de los rasgos no intuitivos de la vida dan las mejores pistas de su origen. Como resultado, han llegado con una imagen radicalmente diferente de lo que era la vida primitiva y donde se desarrolló. Es una imagen para la cual hay cada vez más pruebas.
Antes de Mitchell, todo el mundo suponía que las células obtienen su energía con una química sencilla. La moneda de energía universal de la vida es una molécula llamada ATP. Se la abre y se libera energía. El ATP alimenta la mayor parte de la energía que demandan los procesos en las células, desde la construcción de las proteínas a mover los músculos. A su vez, se pensaba que el ATP se genera a partir de los alimentos por una serie de reacciones químicas estándar. Mitchell pensaba lo contrario. La vida, argumentó, no es impulsada por el tipo de química que ocurre en un tubo de ensayo, sino por una especie de electricidad.
La energía de los alimentos, decía, se usa para bombear iones de hidrógeno con carga positiva, o protones, a través de una membrana. Como los protones se acumulan de un lado, se va acumulando un gradiente electroquímico a través de la membrana. Cuando se presenta la oportunidad, los protones regresan, liberando energía que se aprovecha para ensamblar moléculas de ATP. En términos de energía, el proceso es análogo a llenar un alto tanque con cubos de agua, y luego de usar el agua para que gire una noria.
Mitchell denominó “quimiosmosis” a su teoría, y no es de extrañar que los biólogos la encontraran difícil de aceptar. ¿Por qué la vida va a generar energía de manera tan compleja e indirecta, cuando serían suficientes las reacciones químicas simples? Simplemente no tiene sentido.
Puede ser no intuitiva, pero la quimiosmosis ha resultado omnipresente en el mundo de los seres vivos. La fuerza de los protones mueve no sólo la respiración celular, sino también la fotosíntesis: la energía del Sol se convierte en un gradiente de protones de una manera que es esencialmente la misma que lo que pasa con la energía de los alimentos.
Y los gradientes de protones a menudo se aprovechen directamente, en lugar de ser utilizados para producir ATP. Impulsan la rotación de los flagelos de las bacterias, así como el transporte activo de numerosas sustancias dentro y fuera de las células. Así que el poder de los protones es fundamental para la generación de energía, movimiento, y para mantener el ambiente interno; algunas de las funciones más básicas de la vida.
Esto sugiere que la energía de los protones no es una innovación tardía, sino que evolucionó bien al principio de la historia de la vida, una idea respaldada por el árbol de la vida. La primera rama del árbol se ubica entre los dos grandes grupos de células simples, las bacterias y las arqueas. Ambos grupos tienen bombas de protones, y ambos generan ATP a partir de las corrientes de protones, utilizando una proteína similar. La explicación obvia es que este mecanismo fue heredado de un ancestro común, el progenitor de toda la vida en la Tierra.
Piense acerca de las propiedades de ese ancestro común, dice Bill Martin, de la Universidad de Dusseldorf, en Alemania, y llegará a una bestia muy extraña. Él parte del supuesto de que lo más probable es que las características encontradas tanto en archaea y como en las bacterias sean heredadas de un ancestro común de toda la vida, aunque algunas han sido claramente adquiridas, posteriormente, por un intercambio de genes. Caminos distintos, presumiblemente evolucionados de forma independiente.
No hay duda de que el ancestro común procesaba ADN, ARN y proteínas, tenía un código genético universal, ribosomas (las fábricas de producción de proteínas), ATP y una enzima impulsada por protones para la fabricación del ATP. Los detallados mecanismos para la lectura de ADN y la conversión de los genes en proteínas también estaban allí. En resumen, entonces, el ancestro común de toda la vida se ve muy parecido a una célula moderna.
Y las diferencias son sorprendentes. En particular, el mecanismo detallado de la replicación del ADN habría sido muy diferente. Parece como si la replicación del ADN hubiese evolucionado en forma independiente en bacterias y arqueas, de acuerdo a Eugene Koonin en el Centro Nacional de Información sobre Biotecnología, en Bethesda, Maryland.
Más allá de eso, muchas vías bioquímicas son catalizadas por enzimas muy diferentes. Lo más sorprendente y más importante de esto es la fermentación, la producción de energía a partir de los alimentos sin oxígeno. A menudo se supone que la fermentación es el método primordial de generación de energía. Sin embargo, Martin ha demostrado que las enzimas responsables son totalmente independientes en las archaea y en las bacterias. Parecería como si la fermentación hubiese evolucionado dos veces y más tarde, en lugar de en los albores de la vida.
Límites desconcertantes:
Aún más desconcertante, dice Martin, es que ni las membranas celulares, ni las paredes de las células tengan detalles en común. “A primera vista, los límites que definen a las células evolucionaron de forma independiente en bacterias y en archaea”, dice.
Pero si ese es el caso, ¿qué clase de célula era este ancestro común? ¿Una célula sin paredes? ¡Imposible! ¿Algo único? Si se excluye lo imposible, entonces lo que queda debe ser la verdad.
Si Martín tiene razón, el ancestro común de la vida en la Tierra era una entidad sofisticada en términos de sus genes y proteínas, y se alimentaba con las corrientes de protones en lugar de con la fermentación. Sin embargo, al mismo tiempo, sus membranas eran, al parecer, diferentes a cualquier cosa que exista hoy día. Era vida, pero no como lo conocemos.
Alrededor de 2002 Martín conoció la obra de Russell. Hasta ese momento, Russell había sido una voz solitaria. Sus ideas geoquímicas sobre el origen de la vida no iban bien con los biólogos moleculares que dominaban el campo.
Desde principios de los 90, Russell exploró las posibilidades de un tipo muy particular de ventiladeros hidrotérmicos, a los que se les llama ventiladeros alcalinos, conocidos ahora gracias a los restos que se encuentran en rocas antiguas. A diferencia de las negras fumarolas descubiertas en 1977, formadas por la violenta reacción del agua de mar con lava volcánica que brota de las dorsales oceánicas, los ventiladeros de Russell eran muy mansos, poco más que piedras llenas de burbujas y plagadas de laberínticos poros.
Estos respiraderos se formaron cuando el agua reaccionó con el mineral olivino, común en el fondo del mar, y habrían sido aún más comunes al principio, antes de que la corteza terrestre se espese. El proceso produce un nuevo mineral, llamado serpentina, y libera hidrógeno, fluidos alcalinos y calor. También hace que las rocas se agranden y agrieten, permitiendo que más agua se filtre hacia abajo, lo que mantiene la reacción. Los cálidos efluentes, ricos en hidrógeno, en última instancia rompen a través del fondo del mar como un respiradero hidrotérmico alcalino.
El interés en los respiraderos alcalinos aumentó en 2000, cuando Deborah Kelley y sus colegas de la Universidad de Washington en Seattle tropezaron (si se puede tropezar en un sumergible) con un campo de fumarolas activas alcalinas cerca de la dorsal atlántica, exactamente donde Russell dijo que debían estar esos ventiladeros. El equipo denominó el lugar como “Ciudad Perdida” (Lost City), en parte por las espectaculares torres de roca que se forman cuando los carbonatos se precipitan desde el líquido alcalino.
Al igual que los respiraderos antiguos, las torres de la Ciudad Perdida están plagado de pequeños poros, algunos con dimensiones no muy diferentes a las células modernas. Y la química se ajusta a la teoría, también. Un informe del año pasado confirmó la presencia de metano y otros hidrocarburos pequeños, así como el propio hidrógeno
Los ventiladeros pueden ser los mismos que los de alrededor de 4.000 millones de años atrás, pero en aquel entonces los océanos eran muy diferentes. Los océanos primordiales estaban saturados de dióxido de carbono, lo que los hacía ácidos, mientras que los mares hoy en día son ligeramente alcalinos. Y prácticamente no había oxígeno. Sin oxígeno, el hierro se disuelve fácilmente: las grandes formaciones en de hierro de todo el mundo nos revelan la cantidad de hierro que se disolvió en los océanos. Cuando los niveles de oxígeno crecieron lentamente, miles de millones de toneladas de hierro precipitaron en forma de óxido.
Lo que significa, dice Russell, que la interfaz entre los ventiladeros alcalinos y los antiguos mares habría sido mucho más favorable para la bioquímica primordial de lo que son hoy. En particular, se habrían formado burbujas de minerales de hierro-azufre —que tienen notables propiedades catalíticas— en los poros. Esto no es una conjetura. Russell ha encontrado respiraderos antiguos con una estructura similar, e incluso los reprodujo en el laboratorio.
El hecho de que los respiraderos alcalinos habrían tenido un laberinto de microcompartimientos formados naturalmente es lo que atrajo la atención de Martin. Estos compartimentos podrían haber sido los precursores de las paredes celulares biológicas que él estaba buscando, proporcionando un andamio en el que se podría formar el contenido de las células. Juntos, Russell y Martin han señalado que todavía se pueden encontrar idénticos minerales de hierro-azufre en el núcleo de las proteínas que convierten el dióxido de carbono en azúcares —utilizando gas de hidrógeno— en arqueas y bacterias como los metanógenos y acetógenos.
Los fluidos de los respiraderos contenían compuestos de nitrógeno como el amoniaco, y condiciones que favorecián la producción de aminoácidos, los bloques de construcción de las proteínas.
Eso no es todo. En presencia de fosfato, los minerales podrían haber catalizado la producción de nucleótidos, los bloques de construcción del ARN y ADN. Y si los nucleótidos se formaron por catálisis mineral, los poros en los ventiladeros alcalinos habrían tenido un extraordinario efecto.
Simulaciones por el equipo de Dieter Braun en la Universidad Ludwig Maximilian de Munich, Alemania, muestran que el gradiente de temperatura entre la parte superior e inferior de los poros concentra nucleótidos en un extremo, lo que fomenta que las moléculas se unan para formar cadenas de ARN y ADN (véase el diagrama). Estas moléculas de mayor tamaño pasarían a concentrarse en niveles aún más altos. Es más, las corrientes de convección producirían un continuo aumento y caída de temperatura, algo que se usa ahora para fabricar ADN en laboratorios de todo el mundo.
El mundo de ARN
Los experimentos de laboratorio hechos por un equipo dirigido por el Premio Nobel Jack Szostak de la Universidad de Harvard, publicados a principios de este año, han confirmado que estas condiciones en efecto concentran nucleótidos y ácidos nucleicos. El equipo también encontró que se concentran ácidos grasos, dando lugar a una formación espontánea de burbujas parecidas a células en el interior de los poros.
Es difícil imaginar un mejor escenario para el mundo del ARN que, como se piensa ampliamente, fue el que cerró la brecha entre los productos químicos orgánicos simples y la complejidad del ADN y las proteínas. Así que la idea de que los antiguos respiraderos hidrotermales alcalinos fueron los incubadores de la vida parece muy plausible, incluso antes de considerar su característica más llamativa: un gradiente de protones ya “servido”.
En aquel entonces los mares eran ácidos, y la acidez se define en términos de protones: los ácidos son ricos en ellos. “Los fluidos alcalinos burbujeando en un océano ácido forman células catalíticas minerales “con un gradiente de protones a través de sus membranas inorgánicas”, dice Russell. “Están establecidos en la misma forma peculiar que todas las células actuales”.
Algunos investigadores han rechazado las “células” naturales quimiosmóticas de Russell como una mera curiosidad, irrelevante para el origen de la vida. Pero cuando Martin y Russell examinaron la bioenergética de los más simples procariotas, se dieron cuenta de que las primeras células nunca podrían haber escapado de los ventiladeros sin esa quimiosmosis natural, lo que explica por qué el extraño mecanismo de Mitchell es tan fundamental para la vida actual, y tan universal.
El gran escape.
En toda la increíble riqueza de la vida en la Tierra sólo hay cinco maneras en que se captura el dióxido de carbono y se convierte en materia viva, y sólo uno de ellos no cuesta nada en absoluto. Es la reacción directa de hidrógeno con dióxido de carbono. Esta reacción exotérmica convierte el dióxido de carbono en moléculas orgánicas simples y también libera energía. Se ha descrito como “un almuerzo gratis donde te pagan por comer”. Y si bien el hidrógeno no suele burbujear por sí solo desde la tierra, sí lo hace en los respiraderos alcalinos.
La reacción del hidrógeno con el dióxido de carbono es fundamental para la vida en los respiraderos, pero hay un gran problema: cuesta un poco de energía iniciar la reacción en un principio, mientras que la cantidad de energía necesaria para mantener el crecimiento es insignificante. De hecho, es tan insignificante, de acuerdo con Rolf Thauer en el Max Planck Institute for Terrestrial Microbiology en Marburg, Alemania, que es imposible que las bacterias crecieran sólo por la química: necesitan la energía de la quimiosmosis de protones.
Para entender esto, piense en la energía almacenada por el ATP como un equivalente a 10 dólares. Si toma 10 dólares para poner en marcha una reacción que luego libera 15, en teoría la célula ha ganado 5 dólares. Sin embargo, si la única forma que tiene una célula de almacenar energía es producir ATP, puede hacer sólo una molécula, ya que hacer dos nuevos ATPs costaría 20 dólares. Así que se ha gastado un ATP para obtener un ATP, y los 5 dólares de sobra se desperdiciaron como calor. Eso no es compatible con mantenerse vivo.
Para la proto-vida en los ventiladeros, esto no habría sido problema. El fluido de las chimeneas contenía moléculas reactivas como sulfuro de metilo, lo que generaría acetil fosfato, una molécula que algunas bacterias de hoy todavía usan de manera intercambiable con el ATP. Es más, el gradiente natural de protones complementó esta fuente de energía por generación espontánea de otra forma primitiva del ATP, llamado pirofosfato.
El pirofosfato también actúa de forma muy similar al ATP, y todavía es usado junto con el ATP por muchas bacterias y arqueas. Estas bacterias aceleran su producción utilizando una enzima simple llamada pirofosfatasa. En un trabajo que pronto se publicará realizado junto a Wolfgang Nitschke en el Instituto de Biología Estructural y Microbiología en Marsella, Francia, Russell ha demostrado que esta enzima se encuentra en algunas de las células más primitivas que conocemos y está a ambos lados de la división entre bacterias y arqueas. Lo que sugiere que data bien desde el principio.
Así que el ancestro común de la vida podría haber aprovechado el gradiente natural de protones de los respiraderos antiguos para producir energía. Para escapar de los ventiladeros, sólo era necesario un paso más: revertir el proceso para almacenar energía.
La quimiosmosis permite que las células ahorren pequeñas cantidades de energía, los 5 dólares que de otro modo se perderían. Una reacción que se puede repetir una y otra vez, sólo para bombear protones por una membrana. Es como ahorrar para comprar algo, eventualmente el gradiente de protones será suficiente para producir un pirofosfato o un ATP. El resultado es que los gradientes de protones permiten que las células crezcan, y puedan salir de los ventiladeros.
De hecho, parece que la rareza de Mitchell es una condición previa necesaria para la vida. Si bien la descomposición de azúcares puede proporcionar suficiente energía para crecer sin necesidad de quimiosmosis, el proceso —la glucólisis— involucra complejas vías y requiere montones de azúcar, por lo que es muy improbable que haya sido la fuente principal de energía para la vida inicial.
El cuadro pintado por Russell y Martin es en verdad impresionante. El ancestro común de la vida no era una célula viva independiente, sino una roca porosa llena de burbujas con membrana de sulfuro de hierro que catalizaron las reacciones bioquímicas primordiales con el impulso del hidrógeno y los gradientes de protones, este reactor de flujo natural lleno de productos químicos orgánicos, dando lugar a una proto-vida que finalmente surgió como las primeras células vivientes, no una vez sino dos, dando lugar a las bacterias y las arqueas.
Faltan llenar muchos detalles, y puede que nunca sea posible demostrar más allá de toda duda que la vida evolucionó por este mecanismo. La evidencia, sin embargo, está creciendo. Este escenario coincide con las propiedades conocidas de la vida en la Tierra, es energéticamente plausible, y pone de vuelta a la gran teoría de Mitchell en su legítimo lugar en el centro mismo de la biología.[8]
Expedición a la Ciudad Perdida
en vivo desde “Reacción Rocosa”
--La extraña química que se desarrolla en Ciudad Perdida se debe a su incomparable marco geológico, enraizado en la estructura del propio planeta.
Imaginemos que la Tierra es un melocotón, donde la piel representa la corteza, la carne equivale a la sólida capa de manto subyacente y el hueso corresponde al núcleo de hierro incandescente.
En la dorsal Mesoatlántica, la corteza se está separando lentamente a medida que Norteamerica y Africa se alejan entre sí, a razón de unos 25 milímetros pro año. La separación de la corteza ha dejado expuestas regiones del manto terrestre en el fondo oceánico: el afloramiento de este manto expuesto ha formado el macizo de Atlantis...
Y lo que reviste de mayor interés;en ese momento los fluidos se encuentran muy reducidos (en el sentido químico), es decir, que el agua ha sido despojada de todo su oxígeno para ser sustituido por gases de alto contenido energético, como el hidrógeno, el metano y el sulfuro.
En concreto, la concetración de hidrógeno es de las más elevadas que se han hallado nunca en un entorno natural.
Por último, la serpentinización da lugar a condiciones químicas que posibilitan la síntesis de compuestos orgánicos a partir de sustancias inorgánicas, un prerequisito para la aparición de la vida. Cabe la posibilidad de que, hace miles de millones de años, fumarolas similares hubieran reunido las condiciones apropiadas para el desarrollo en parte, de la vida en la Tierra.
El extraño campo de ventilas hidrotermales descubierto, sorprendió a los científicos no solamente por sus ventilas, que son las más altas vistas hasta ahora (la que tiene 18 pisos de alto empequeñece a la mayoría de las ventilas de otros sitios, por al menos cien pies). sino también porque los fluidos que forman esas ventilas son calentados por el agua del mar reaccionando con rocas del manto de millones de años de edad, y no por un vulcanismo joven.
...
El extraordinario campo de ventilas hidrotermales “Ciudad Perdida” (Lost City), fue llamado así en parte porque se encuentra en una montaña del suelo marino llamada Macizo Atlantis, fue descubierto en el medio del océano Atlántico a unas 1.500 millas de la Costa Este de los EE.UU.: durante una expedición que ni siquiera estaba buscando ventilas hidrotermales.
...
Vida Eruptiva, Pero No Volcánica
La Ciudad Perdida es distinta en parte porque la poderosa ventila de 180 pies del sitio, a la que los científicos llaman Poseidón, es mucho más grande que las ventilas de humo negro estudiadas previamente, que en su mayoría miden 80 pies o menos. La más alta chimenea de humo negro vista jamás era una ventila de 135 pies (que se ha derrumbado en años recientes) en las afueras de la costa de Washington.
Contrastando con las ventilas de humo negro, que son una oscura mezcla moteada de minerales sulfhídricos, las ventilas de la Ciudad Perdida son casi 100 por ciento carbonato, el mismo material que la piedra caliza de las cavernas, y varían en color desde un hermoso blanco limpio hasta el crema o gris.
Las diferencias son causadas por la ventilación hidrotermal (un proceso en el cual el agua circula hacia dentro del fondo marino, adquiriendo calor y productos químicos hasta que los fluidos están lo suficientemente calientes como para retornar al océano), no parece estar conectada con la actividad volcánica y las cámaras de magma. Ésto es diferente a la mayoría de los sistemas de los centros de expansión de la cordillera centro-oceánica. Allí es donde un joven suelo marino es creado, a menudo dramáticamente, durante las erupciones volcánicas, y el agua de las ventilas puede llegar a los 700 F.
La Vida Diaria en la Ciudad Perdida
Los fluidos de ventila de la Ciudad Perdida mantienen una comunidad de microorganismos que se cree viven de los gases metano e hidrógeno, ambos sub-productos de la serpentinización. Ésto lleva a que Kelley, Karson y otros especulen que la vida en este planea podría haber comenzado precisamente en un ambiente como éste, dado que, principalmente, mucha más roca del manto fue expuesta al agua de mar muy temprano en la historia de la Tierra. Y lo mismo pudo haber sucedido en otros mundos.
Biosferas ocultas de la Tierra
Dos recientes descubrimientos en el campo de la astrobiología desafían nuestros principales supuestos acerca de una comunidad biológica integrada en la Tierra. A nivel microbiano, que podría haber biosferas ocultas existiendo en la Tierra junto con nuestros vecinos más conocidos. Una comunidad de este tipo ha sido encontrada enterrada profundamente, mientras que la otra vive en el mar junto con otras formas de vida más conocidas.
Potencialmente, entre los descubrimientos recientes más importantes para la astrobiología está el hallazgo de formas de vida profundamente enterradas que parecen desarrollarse independientemente de la biosfera superficial conocida, que está animada por la luz solar. Estos microorganismos, descubiertos en agua subterránea caliente a 2.8 km de profundidad en una mina de oro de Sudáfrica, obtienen su energía en última instancia a partir de compuestos químicos –hidrógeno y sulfatos- producidos por un lento decaimiento de elementos radioactivos en las rocas. La existencia de una comunidad microbiana profundamente subterránea en la Tierra plantea que podrían existir biosferas aisladas semejantes en otros planetas, como Marte, a pesar de las condiciones hostiles reinantes en sus superficies.
Un importante equipo internacional de investigación, liderado por científicos de las Universidades de Princeton e Indiana y respaldado en parte por el Instituto de Astrobiología de la NASA, anunció su descubrimiento en una monografía publicada en la edición del 20 de Octubre de 2006 de la revista Science. El autor principal de la monografía es Li-Hing Lin, perteneciente en este momento a la Universidad Nacional de Taiwán, pero que comenzó su trabajo como estudiante graduado de Princeton. Los científicos aprovecharon una fractura inundada de agua cruzada durante una perforación en una profunda mina de oro cerca de Johannesburgo. Comenzaron a recolectar muestras de agua a los pocos días de flujo inicial para asegurar una mínima contaminación, y continuaron extrayendo nuevas muestras durante varios meses.
Utilizando modernas herramientas de análisis genético, compararon los microorganismos con otras comunidades microbianas anaeróbicas que obtienen su energía a partir de la reducción de sulfato. Un estudio detallado de la composición química del agua indica que hay suficiente sulfato e hidrógeno de origen natural como para sostener la vida indefinidamente. La base de la cadena alimentaria es un reductor de sulfato que pertenece al filum llamado Firmicutes, y los otros microbios en la comunidad pueden subsistir gracias a los productos que obtienen a partir de su productor primario. El agua misma fue datada en decenas de millones de años, un tiempo durante el cual no ha tenido contactos físicos o químicos con el mundo de más arriba, más conocido.
En los últimos años, los científicos han descubierto muchas comunidades de microorganismos que viven bajo condiciones que antes se consideraban imposibles para la vida. Los extremófilos pueden prosperar bajo condiciones de temperaturas muy altas o muy bajas, o una composición química ácida o alcalina, o una elevada salinidad que mataría a microorganismos más conocidos de la superficie. Algunos de estos extremófilos viven en comunidades en respiraderos hidrotermales en las profundidades del océano o dentro de rocas, a cientos de metros bajo la superficie. Sin embargo, hasta ahora la “biosfera profunda” parecía necesitar, por lo menos indirectamente, un contacto con la atmósfera. Este descubrimiento, hecho en un ambiente mucho más profundo y aislado, representa un avance fundamental, revelando una forma de vida alienígena dentro de la Tierra.
Este descubrimiento puede proporcionar indicios claves sobre la habitabilidad de Marte y otros planetas con superficies hostiles pero agua caliente en el interior. Las misiones de la NASA revelaron que Marte alguna vez tuvo un ambiente superficial más apto para la vida, con agua líquida y luz solar, circunstancias bajo las cuales la vida puede haberse formado y prosperado hace miles de millones de años. Quizás el remanente de aquella vida persista bajo su superfice, en ambientes como este mar sin sol en la Tierra.
El segundo descubrimiento excitante es el de una hasta ahora desconocida “biosfera rara” que coexiste con formas de vida más conocidas en lo profundo del océano. Los descubridores son un equipo de científicos del Laboratorio Biológico Marino de Woods Hole y el Real Instituto Holandés para la Investigación Marina, bajo la conducción de Mitch Sogin, quien también es el PI para el equipo del MBL en el NAI. Los científicos utilizaron nuevas herramientas de análisis genético para buscar las poblaciones microbianas dominantes para extraer muestras de los microbios mucho más raros que antes no habían sido detectados, usando muestras recogidas de agua marina normal y el agua caliente asociada con los respiraderos hidrotermales. Su trabajo fue publicado en los Proceedings de la Academia Nacional de Ciencias del 8 de agosto de 2006.
Este nuevo análisis revela una importante diversidad dentro de esta “rar biosfera”. Las técnicas utilizadas, si bien no permite el aislamiento de organismos individuales para su estudio, sí posibilitan la elaboración de cálculos estadísticos de la población. Pero aunque el número de tales microorganismos es pequeño, hay por lo menos una diversidad de especies 100 veces mayor de lo que se preveía.
¿Qué están haciendo estos diversos organismos? En cierto modo, no son muy exitosos, dado que son superados en número ampliamente por especies más conocidas. Sin embargo, ellos subsisten en pequeños números. Quizás representen una clase de “sistema de seguridad” natural o fuerza de reserva que puede entrar en acción si las condiciones ambientales cambian de manera tal que amenace al ecosistema dominante.
Podrían ser parte de la estrategia de la vida para sobrevivir ante cambios ambientales catastróficos. El próximo desafío es calcular los patrones globales de esta “rara biosfera” y comenzar a caracterizar a sus especies individuales. [9]
El carbono es un elemento químico de número atómico 6 y símbolo C. Es sólido a temperatura ambiente. Dependiendo de las condiciones de formación, puede encontrarse en la naturaleza en distintas formas alotrópicas, carbono amorfo y cristalino en forma de grafito o diamante. Es el pilar básico de la química orgánica; se conocen cerca de 15 millones de compuestos de carbono, aumentando este número en unos 500.000 compuestos por año, y forma parte de todos los seres vivos conocidos. Forma el 0,2 % de la corteza terrestre.
[10]
La química orgánica se constituyó como disciplina en los años treinta. El desarrollo de nuevos métodos de análisis de las sustancias de origen animal y vegetal, basados en el empleo de disolventes como el éter o el alcohol, permitió el aislamiento de un gran número de sustancias orgánicas que recibieron el nombre de "principios inmediatos". La aparición de la química orgánica se asocia a menudo al descubrimiento, en 1828, por el químico alemán Friedrich Wöhler, de que la sustancia inorgánica cianato de amonio podía convertirse en urea, una sustancia orgánica que se encuentra en la orina de muchos animales. Antes de este descubrimiento, los químicos creían que para sintetizar sustancias orgánicas, era necesaria la intervención de lo que llamaban ‘la fuerza vital’, es decir, los organismos vivos. El experimento de Wöhler rompió la barrera entre sustancias orgánicas e inorgánicas. Los químicos modernos consideran compuestos orgánicos a aquellos que contienen carbono e hidrógeno, y otros elementos (que pueden ser uno o más), siendo los más comunes: oxígeno, nitrógeno, azufre y los halógenos. Por ello, en la actualidad, la química orgánica tiende a denominarse química del carbono.[11]
La química inorgánica se encarga del estudio integrado de la formación, composición, estructura y reacciones quimicas de los elementos y compuestos inorgánicos (por ejemplo, ácido sulfúrico o carbonato cálcico); es decir, los que no poseen enlaces carbono-hidrógeno, porque éstos pertenecen al campo de la química orgánica. Dicha separación no es siempre clara, como por ejemplo en la química organometálica que es una superposición de ambas.
El nombre tiene su origen en la época en la que todos los compuestos del carbono se obtenían de seres vivos; de ahí la química del carbono se denomina química orgánica. La química de compuestos sin carbono, fue, por ende, llamada química inorgánica.[12]
Oxidaciones y reducciones biológicas
El metabolismo implica cientos de reacciones redox. Así, el catabolismo lo constituyen reacciones en que los sustratos se oxidan y las coenzimas se reducen. Por el contrario, las reacciones del anabolismo son reacciones en que los sustratos se reducen y los coenzimas se oxidan. En su conjunto, catabolismo y anabolismo constituyen el metabolismo.[13]
El trifosfato de adenosina o adenosín trifosfato (ATP, del inglés adenosine triphosphate) es un nucleótido fundamental en la obtención de energía celular. Está formado por una base nitrogenada (adenina) unida al carbono 1 de un azúcar de tipo pentosa, la ribosa, que en su carbono 5 tiene enlazados tres grupos fosfato. Se encuentra incorporada en los ácidos nucleicos.
ATP y metabolismo
El acoplamiento entre las reacciones exergónicas que liberan energía al medio y endergónicas (con consumo de energía), en conjunto constituyen el metabolismo celular.
Las reacciones endergónicas se manifiestan durante los procesos anabólicos; de manera que, requieren que se le añade energía a los reactivos (sustratos o combustibles metabólicos), se le suma energía (contiene más energía libre que los reactivos). Por otro lado, durante las reacciones exergónicas se libera energía como resultado de los procesos químicos (ej, el catabolismo de macromoléculas). La energía libre se encuentra en un estado organizado, disponible para trabajo biológico útil. Las reacciones endergónicas se llevan a cabo con la energía liberada por las reacciones exergónicas. Las reacciones exergónicas pueden estar acopladas con reacciones endergónicas. Reacciones de oxidación-reducción (redox) son ejemplos de reacciones exergónicas y endergónicas acopladas.[14]
Los organismos pluricelulares nos alimentamos principalmente de metabolitos complejos (proteínas, lípidos, glúcidos) que degradamos a lo largo del tracto intestinal, de modo que a las células llegan metabolitos menos complejos que los ingeridos.
En la célula son oxidados por una serie de reacciones químicas degradativas (catabolismo). Como productos del catabolismo se obtienen metabolitos simples y energía. Ambos son los precursores para la síntesis de los componentes celulares. Todo el conjunto de reacciones de síntesis se llama anabolismo. En el catabolismo (oxidación) se produce una liberación de electrones que son captados por moléculas transportadoras de electrones como el NAD+ (que al aceptar electrones se reduce a NADH).
Por otra parte, la energía liberada queda retenida en su mayoría en el ATP.
La síntesis (anabolismo) de los compuestos celulares se realiza con los metabolitos simples, utilizando la energía contenida en el ATP y los electrones contenidos en el NADH, ya que éste es un proceso reductivo (toma electrones). El ATP es esa moneda de intercambio energético debido a su estructura química. Cuando se hidroliza libera mucha energía que va a ser captada por las enzimas que catalizan las reacciones de biosíntesis.[15]
Las biomoléculas son las moléculas constituyentes de los seres vivos. Los cuatro bioelementos más abundantes en los seres vivos son el carbono, hidrógeno, oxígeno y nitrógeno, representando alrededor del 99% de la masa de la mayoría de las células. Estos cuatro elementos son los principales componentes de las biomoléculas debido a que:
Permiten la formación de enlaces covalentes entre ellos, compartiendo electrones, debido a su pequeña diferencia de electronegatividad. Estos enlaces son muy estables, la fuerza de enlace es directamente proporcional a las masas de los átomos unidos.
Permiten a los átomos de carbono la posibilidad de formar esqueletos tridimensionales –C-C-C- para formar compuestos con número variable de carbonos.
Permiten la formación de enlaces múltiples (dobles y triples) entre C y C, C y O, C y N, así como estructuras lineales ramificadas cíclicas, heterocíclicas, etc.
Permiten la posibilidad de que con pocos elementos se den una enorme variedad de grupos funcionales (alcoholes, aldehídos, cetonas, ácidos, aminas, etc.) con propiedades químicas y físicas diferentes.
Son sintetizadas solamente por los seres vivos y tienen una estructura a base de carbono. Están constituidas principalmente por carbono, hidrógeno y oxígeno, y con frecuencia están también presentes nitrógeno, fósforo y azufre; otros elementos son a veces incorporados pero en mucha menor proporción.
Las biomoléculas orgánicas pueden agruparse en cuatro grandes tipos: Glúcidos, proteinas, lípidos y ácido nucleicos
Los glúcidos (hidratos de carbono o carbohidratos) son la fuente de energía primaria que utilizan los seres vivos para realizar sus funciones vitales; la glucosa está al principio de una de las rutas metabólicas productoras de energía más antigua, la glucólisis, usada en todos los niveles evolutivos, desde las bacterias a los vertebrados. Muchos organismos, especialmente los de estirpe vegetal (algas, plantas) almacenan sus reservas en forma de almidón. Algunos glúcidos forman importantes estructuras esqueléticas, como la celulosa, constituyente de la pared celular vegetal, o la quitina, que forma la cutícula de los artrópodos.[8]
La glucólisis o glicolisis (del griego glycos: azúcar y lysis: ruptura), es la vía metabólica encargada de oxidar o fermentar la glucosa y así obtener energía para la célula. Ésta consiste en 10 reacciones enzimáticas que convierten a la glucosa en dos moléculas de piruvato, la cual es capaz de seguir otras vías metabólicas y así continuar entregando energía al organismo.
El complejo ATP sintasa también denominada complejo V o FoF1-ATP-sintasa (F = factor de acoplamiento, en inglés coupling factor) y también conocida como ATP-asa, por su funcionamiento, hidróliza el ATP. Es una enzima situada en la cara interna de la membrana interna de las mitocondrias y de la membrana de los tilacoides de los cloroplastos encargada de sintetizar ATP a partir de ADP y un grupo fosfato y la energía suministrada por un chorro proton (H+). Responde a la síntesis de ATP según la hipótesis quimiosmótica de Mitchell. La síntesis de ATP gracias a este enzima se denomina del ADP. La ATP sintasa se puede observar como un motor molecular que produce una gran cantidad de ATP cuando los protones fluyen a través de ella. La tasa de sintesis es grande, el organismo humano en fase de reposo puede formar unas 1021 moléculas de ATP por segundo.[16]
La Hipótesis quimiosmótica también conocida como la hipótesis quimiosmótica de Mitchell, explica la manera en que la energía libre que se genera por la actividad de cadena de transporte de electrones se emplea para producir ATP a partir de la reacción ADP + Pi. Las coenzimas reducidas procedentes del Ciclo de Krebs (NADH + H+ y FADH2) atraviesan una serie de complejos enzimáticos (cadena de transporte de electrones) que producen en ellas sucesivas reacciones de oxidación-reducción cuyo objetivo es la utilización del flujo de electrones para bombear protones al espacio intermembrana de la mitocondria o cloroplasto, creando un gradiente electroquímico un gradiente de pH (gradiente de concentración) el y de un desequilibrio de carga (gradiente electroquímico) en la membrana mitocondrial interna. Las ATP sintase forman ATP utilizando la energía libre procedente del gradiente de protones, utilizando el hecho de que los protones tienden a volver al interior del orgánulo, e sólo se pueden hacer a través de la ATP sintasa, por el hecho de que la membrana interna es impermeable a los protones. Cuando los protones son bombeados fuera de la matriz, el espacio intermembrana se convierte en más ácido y con mayor carga positiva que en la matriz. La matriz es básica y con una carga negativa. Peter Denis Mitchell (1920-1992) utilizó el término “quimiosmótico” para describir las reacciones enzimáticas en las que intervienen, simultáneamente, una reacción química y un proceso de transporte.
Acoplamiento del transporte electrónico y la síntesis de ATP
El gradiente electroquímico acopla el ritmo de la cadena de transporte electrónico con el ritmo de la síntesis de ATP. Debido a que el flujo electrónico necesita el bombeo de protones, el flujo electronico no puede producirse más rápidamente que la utilización de los protones para síntesis de ATP ( fosforilación oxidativa acoplada), significando en una relación estrechamente acoplada entre la oxidación y la fosforilación. Esto conlleva a que los sustratos se oxidan, los electrones se transportan y el oxígeno se consumo tan sólo cuando se requiere la síntesis de ATP. Por lo tanto, las mitocondrias en reposo consumen oxígeno a una velocidad lenta, pero que puede incrementar enormemente en la presencia de ADP. El ADP es captado por las mitocondrias y estimula la ATP sintasa, que disminuye el gradiente de protones. Entonces aumenta la respiración, puesto que son estimuladas las bombas de protones para restablecer el gradiente. Por tanto se puede resumir diciendo que el “control respiratorio” es la dependencia de captación de oxígeno por las mitocondrias según la disponibilidad de ATP.
Transporte de ATP y ADP
La fuerza protón-motriz también sirve para impulsar varios procesos de transporte esenciales para la fosforilación oxidativa, aunque el papel principal del gradiente de protones en las mitocondrias es suministrar energía para la síntesis de ATP. La membrana mitocondrial interior necesita transportadores especializados para el ADP y Pi desde el citosol hacia el interior de la mitocondria, al sitio donde se puede resíntetizar a ATP, ya que la membrana mitocondrial interna es generalmente impermeable a las moléculas con carga. Por tanto hay dos sistemas específicos que transportan ADP y Pi a la matriz y ATP hacia el citosol externo. Un transportador de nucleótidos de adenina translocasa y fosfato translocasa son los dos sitemas que intervienen.[4] La nucleótido de adenina translocasa une ADP y es por tanto la encargada en transportar una molécula de ADP desde el citosol hacia el interior de la mitocondria, a la vez que exporta un ATP desde la matriz de nuevo hacia el citosol. Fosfato translocasa transporta en Pi junto con un protón a la matriz.[17]
La cadena de transporte de electrones es una serie de transportadores de electrones que se encuentran en la membrana plasmática de bacterias, en la membrana interna mitocondrial o en las membranas tilacoidales, que median reacciones bioquímicas que producen adenosina trifosfato (ATP), que es el compuesto energético que utilizan los seres vivos. Sólo dos fuentes de energía son utilizadas por los organismos vivos: reacciones de óxido-reducción (redox) y la luz solar (fotosíntesis).
En esta etapa se oxidan las coenzimas reducidas, el NADH se convierte en NAD+ y el FADH2 en FAD+. Al producirse esta reacción, los átomos de hidrógeno (o electrones equivalentes), son conducidos a través de la cadena respiratoria por un grupo de transportadores de electrones, llamados citocromos. Los citocromos experimentan sucesivas oxidaciones y reducciones (reacciones en las cuales los electrones son transferidos de un dador de electrones a un aceptor).
En consecuencia, en esta etapa final de la respiración, estos electrones de alto nivel energético descienden paso a paso hasta el bajo nivel energético del oxígeno (último aceptor de la cadena), formándose de esta manera agua.
Los organismos que utilizan las reacciones redox para producir ATP se les conoce con el nombre de quimioautótrofos, mientras que los que utilizan la luz solar para tal evento se les conoce por el nombre de fotoautótrofos. Ambos tipos de organismos utilizan sus cadenas de transporte de electrones para convertir la energía en ATP.[18]
-HIPÓTESIS QUIMIOSMÓTICA
Durante mucho tiempo se intentó explicar la naturaleza del enlace entre la cadena respiratoria y el sistema de fosforilación. En 1961, Mitchell propuso la hipótesis quimiosmótica, que es la que actualmente se acepta en general.
Esta hipótesis ha sido apoyada por las evidencias experimentales encontradas en distintos laboratorios, lo que le valió a Mitchell el premio Nobel en 1978.
La misma propone que el transporte de electrones y la síntesis de ATP están acopladas por un gradiente protónico a través de la membrana mitocondrial.
Según este modelo, el transporte de electrones paso a paso, desde el NADH o el FADH2 hasta el oxígeno a través de los transportadores de electrones, da por resultado el bombeo de protones a través de la membrana mitocondrial interna hacia el espacio entre las membranas mitocondriales interna y externa.
Este proceso genera un potencial de membrana a través de la membrana mitocondrial interna, ya que el medio que ocupa el espacio intermembranoso se carga positivamente.
La diferencia en concentración de protones entre la matriz y el espacio intermembranoso representa energía potencial, resultado en parte de la diferencia de pH y en parte de la diferencia en la carga eléctrica de los lados de la membrana. Cuando los protones pueden fluir de regreso a la matriz, descendiendo por el gradiente protónico, se libera energía utilizable en la síntesis de ATP a partir de ADP y Pi.
Los protones regresan a la matriz a través de conductos especiales situados en la membrana interna. Estos conductos están dados por un gran complejo enzimático, llamado ATP SINTETASA. Este complejo consta de dos proteínas: F0 y F1.
Las partículas F0 están incluidas en la membrana mitocondrial interna y la atraviesan desde afuera hacia adentro. Se presume que poseen un conducto o poro interior que permite el paso de los protones. Las partículas F1 (que ya habíamos mencionado, al describir la estructura mitocondrial) son proteínas globulares grandes consistentes en nueve subunidades polipeptídicas unidas a las partículas F0 en el lado de la membrana que linda con la matriz. Se comprobó que propulsa la síntesis de ATP a partir de ADP y Pi. Conforme los protones descienden a lo largo del gradiente de energía, dicha energía utiliza para sintetizar ATP. De esta manera, el gradiente protónico que existe a través de la membrana mitocondrial interna acopla la fosforilación con la oxidación.
RESUMEN
En el afán de analizar detenidamente cada paso de las reacciones metabólicas de fotosíntesis y respiración, perdemos la noción de estos procesos globalmente.
En la fotosíntesis, la energía lumínica se convierte en química y se fija carbono en compuestos orgánicos.
Los fotosintetizadores o autótrofos elaboran hidratos de carbono a partir de CO2 y agua y liberan O2 a la atmósfera. Son estos organismos los que mantienen estables las concentraciones de CO2, y O2 atmosféricos.
En la respiración aeróbica los compuestos orgánicos son degradados a CO2 y H2O con la concomitante producción de energía química bajo la forma de ATP.
http://www.virtual.unal.edu.co/cursos/c ... _07_02.jpg
Al igual que la fosforilación oxidativa en las mitocondrias, la fotofosforilación en los cloroplastos es un proceso de acoplamiento quimiosmótico.[19]
FOTOSÍNTESIS (de φῶς, φωτός luz, σύν juntamente y θέσις colocación): formación de un compuesto químico o una sustancia orgánica mediante la luz.[23]
La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos:
La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis; luego irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser transformada en materia propia por los diferentes seres vivos.
Produce la transformación de la energía luminosa en energía química, necesaria y utilizada por los seres vivos
En la fotosíntesis se libera oxígeno, que será utilizado en la respiración aerobia como oxidante.
La fotosíntesis fue causante del cambio producido en la atmósfera primitiva, que era anaerobia y reductora.
De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis.
Se puede deducir que la diversidad de la vida existente en la Tierra depende principalmente de la fotosíntesis. [22]
La fotosíntesis (del griego antiguo φώτο [foto], "luz", y σύνθεσις [síntesis], "unión") es la conversión de energía luminosa en energía química estable, siendo el adenosín trifosfato (ATP) la primera molécula en la que queda almacenada esa energía química. Con posterioridad, el ATP se usa para sintetizar moléculas orgánicas de mayor estabilidad. Además, se debe de tener en cuenta que la vida en nuestro planeta se mantiene fundamentalmente gracias a la fotosíntesis que realizan las algas, en el medio acuático, y las plantas, en el medio terrestre.[24]
KillBastardsII escribió:Para ir acabando estas rondas de "quotes" de esta primera parte del mensaje(el original no me cabía en un sólo post por pasar los 100000 carácteres, he tenido que dividirlo en dos, enseguida que pueda, colgaré la segunda parte de este mensaje) expondré otras válidas definiciones etimológicas y globales del concepto: "fotosíntesis"


KillBastardsII escribió:La definición que das encajaría si vieramos el proceso de fotosintesis desde una perspectiva botánica. Si usarais una perspectiva general e información más veraz sabrías que la fotosintesis es un proceso de: "disociación de moléculas orgánicas complejas por efecto de la luz, y se define como la interacción de una o más fotones con una molécula objetivo."
Esta y no otra es una definición general de la fotosintesis que es lo que para el tema en cuestión nos interesa. Tú definición de fotosíntesis es muy pobre vista desde esta perspectiva general.
Las ideas que se pueden refrendar observando la fotosíntesis de manera multidimensional son que siempre que hay fotosíntesis hay un transporte gradoelectrónico
.Killbastards escribió:Bajo estas y otras premisas podemos a llegar a comprender que en el universo existen 2 tipos de electromagnetismo, el electromagnetimo centrípato y el centrífugo. El electromagnetismo centrípato tiene que ver con las correlaciones cuanticas propias y el electromagnetismo centrífugo tiene que ver con las correlaciones cuánticas generales.
Moki_X escribió:Joder... hasta aqui he llegado a leer.
Bajo esta y otras premisas... ¿Cuales son esas otras premisas?
Define electromagnetismo centrípato y centrífugo. Porque no lo has definido ni has dicho en qué consiste.
Ansioso espero a que me respondas.
![[qmparto]](/images/smilies/net_quemeparto.gif "Que me parto!")
Bajo esta y otras premisas... ¿Cuales son esas otras premisas?
Define electromagnetismo centrípato y centrífugo. Porque no lo has definido ni has dicho en qué consiste.
Kyubi chan escribió:killbastards tio, que yo no hago la fotosíntesis, y mi perro tampoco. No le des más vueltas.
Di que te has equivocado y no pasa nada, se puede seguir debatiendo.
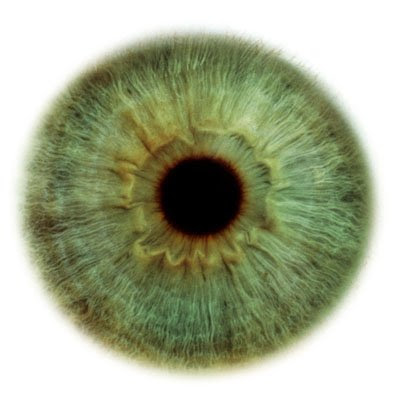
La visión es un sentido que consiste en la habilidad de detectar la luz y de interpretarla (ver). La visión es propia de los animales teniendo éstos un sistema dedicado a ella llamado sistema visual. La visión artificial extiende la visión a las máquinas.
Las células de la retina forman el sistema sensorial del ojo. Las primeras en intervenir son los fotorreceptores, los cuales capturan la luz que incide sobre ellos. Sus dos tipos son los conos y los bastones. Otras células de la retina se encargan de transformar dicha luz en impulsos electroquímicos y en transportarlos hasta el nervio óptico. Desde allí, se proyectan a importantes regiones como el núcleo geniculado lateral y la corteza visual del cerebro.
Las células de la retina forman el sistema sensorial del ojo. Las primeras en intervenir son los fotorreceptores, los cuales capturan la luz que incide sobre ellos. Sus dos tipos son los conos y los bastones. Otras células de la retina se encargan de transformar dicha luz en impulsos electroquímicos y en transportarlos hasta el nervio óptico. Desde allí, se proyectan a importantes regiones como el núcleo geniculado lateral y la corteza visual del cerebro.
En el cerebro comienza el proceso de reconstruir las distancias, colores, movimientos y formas de los objetos que nos rodean.
Los compuestos fotosensibles en la mayoría de los animales así como en los humanos se componen de una proteína llamada opsina, y retineno-1 que es un aldehído de la Vitamina A1.
La Rodopsina es el pigmanto fotosensible de los bastones, cuya opsina se llama escotopsina. La rodopsina capta luz con una sensibilidad máxima en los 505 nm de longitud de onda, esta luz incidente hace que la rodopsina cambie su conformación estructural, produciendo una cascada de reacciones que amplifican la señal, y crean un potencial de acción que se desplazará a través de las fibras nerviosas, y que el cerebro interpretará como luz.[25]
El ojo es un órgano que detecta la luz, por lo que es la base del sentido de la vista.
Se compone de un sistema sensible a los cambios de luz, capaz de transformar éstos en impulsos eléctricos. Los ojos más sencillos no hacen más que detectar si los alrededores están iluminados u oscuros. Los más complejos sirven para proporcionar el sentido de la vista.
Los ojos compuestos se encuentran en los artrópodos (insectos, arácnidos, miriápodos, crustáceos, etc.) y están formados por muchas facetas simples llamadas omatidios que dan una imagen en mosaico (no imágenes múltiples, como a menudo se cree).
En la mayoría de los vertebrados y algunos moluscos, el ojo funciona como una cámara, proyectando imágenes en la retina, donde la luz se transforma gracias a unas células llamadas fotorreceptoras en impulsos nerviosos que son trasladados a través del nervio óptico al cerebro.[26]
El cristalino es un componente del ojo con forma de lente biconvexa que está situado tras el iris y delante del humor vítreo. Su propósito principal consiste en permitir enfocar objetos situados a diferentes distancias. Este objetivo se consigue mediante un aumento de su curvatura y de su espesor, proceso que se denomina acomodación. El cristalino se caracteriza por su alta concentración en proteínas, que le confieren un índice de refracción más elevado que los fluidos que lo rodean. Este hecho es el que le otorga su capacidad para refractar la luz, ayudando a la córnea a formar las imágenes sobre la retina.
Metabolismo del cristalino
La principal fuente de energía del cristalino es la glucosa procedente del humor acuoso. La glucosa entra en las células mediante transportadores independientes de insulina localizados en la membrana plasmática.[6] Cerca del 80% de la glucosa se consume en el cristalino utilizando la glicolisis anaerobia. La utilización aerobia de la glucosa en el ciclo del ácido cítrico se realiza únicamente en las células del epitelio, ya que éstas son las únicas que conservan mitocondrias
La síntesis de proteínas nuevas se detiene cuando las células se transforman en fibras. Todos los cambios que se producen a partir de ese momento son modificaciones post-traduccionales. Por ejemplo, se produce la fosforilación de muchos tipos de proteínas, como cristalinas, proteínas del citoesqueleto y MIP26. Además, se han detectado proteínas con actividad proteasa en el cristalino [27]
La retina de los vertebrados es un tejido sensible a la luz situado en la superficie interior del ojo. Es similar a una tela donde se proyectan las imágenes. La luz que incide en la retina desencadena una serie de fenómenos químicos y eléctricos que finalmente se traducen en impulsos nerviosos que son enviadas hacia el cerebro por el nervio óptico.
La retina tiene una estructura compleja. Esta formada basicamente por varias capas de neuronas interconectadas mediante sinapsis. Las únicas células sensibles directamente a la luz son los conos y los bastones. Los bastones funcionan principalmente en condiciones de baja luminosidad y proporcionan la visión en blanco y negro, los conos sin embargo están adaptados a las situaciones de mucha luminosidad y proporcionan la visión en color.
Células de la retina
La retina tiene tres tipos de células:
-Pigmentadas: Se encargan del metabolismo de los fotorreceptores.
-Neuronas: Hay seis tipos:
:Células fotorreceptoras: Son los conos y los bastones. Los conos son células fotorreceptoras que nos permiten ver los colores, los bastones son celulas fotorreceptoras que nos permiten ver en tonalidades grisáceas cuando existe poca luz.
:Células horizontales.
:Células bipolares de la retina.
:Células amacrinas.(Horizontales, bipolares y amacrinas son neuronas integradoras.)
:Células interplexiformes.
:Células ganglionares de la retina.
-Células de sostén:
:Astrocitos.
:Células de Müller.
Algunos hitos en el estudio de la retina:
A fines del siglo XIX e inicios del siglo XX el español Santiago Ramón y Cajal hizo cortes histológicos de la retina y observándoles en la platina de un microscopio óptico descubrió las células bipolares llamadas conos y bastones, los conos están especializados cada uno para ser sensibles a los colores (basicamente el verde, rojo y azul) los bastones a la baja densidad de luz. En 1952 el alemán Stephen Kuffler estudiando la visión nocturna de los gatos registró la presencia y actividad de células ganglionares que reaccionan individualmente a los estímulos luminosos e incluso a la falta de luz. En el 2007 King-Wai Yau detectó la presencia de células ganglionares planas en las retinas de peces. Es de notar que muchos animales de hábitos nocturnos (felinos, canidos, peces predadores como los tiburones, peces abísales etc.) poseen membranas llamadas "caperuzas" o tapetums tras las retinas, estas membranas funcionan como un espejo que refleja hacia la cornea la luz "excedente" que ha pasado a través de la retina, tal luz reflejada a la cornea es repercibida por la retina (al pasar dos veces por la misma) de estos animales potenciándose en ellos la visión en la oscuridad (nictalopía).[28]
Los bastones son células fotorreceptoras de la retina responsables de la visión en condiciones de baja luminosidad. Presentan una elevada sensibilidad a la luz aunque se saturan en condiciones de mucha luz y no detectan los colores. Se ubican en casi toda la retina exceptuando la fóvea. Contienen rodopsina, que es una proteína que presenta una mayor sensibilidad a las longitudes de onda cercanas a 500 nanometros, es decir, a la luz verde azulada.
Estas células son muy sensibles, capaces de detectar la energía de un sólo fotón y las responsables por tanto de que sea posible la visión nocturna.[29]
Rodopsina
Se encuentra en los discos de los bastones. Es una proteína transmembranal (entre membrana). Consta de una parte proteica, opsina, y una no proteica que es un derivado de la vitamina A que es el 11-cis-retinal.
La opsina es una cadena polipeptídica formada por unos 348 aminoácidos. La opsina se distribuye en siete tramos de hélice alfa que se sitúan perpendiculares a la membrana unidos por partes proteicas sin estructura.
El carboxilo terminal se sitúa en la parte citosólica y el amino en posición intradiscal.
El 11-cis-retinal se sitúa unido a una de las hélices alfa en el centro de la molécula y colocado perpendicularmente. Esta colocación hace que cuando llegue luz incida en el 11-cis-retinal y este se transforma produciendo reacciones que llevan a un impulso nervioso.[30]
Hélice alfa
Los aminoácidos en una hélice α están dispuestos en una estructura helicoidal dextrógira, con unos 3.6 aminoácidos por vuelta. Cada aminoácido supone un giro de unos 100° en la hélice, y los Cα de dos aminoácidos contiguos están separados por 1.5Å. La hélice está estrechamente empaquetada; de forma que no hay casi espacio libre dentro de la hélice. Todas las cadenas laterales de los aminoácidos están dispuestas hacia el exterior de la hélice.
El grupo N-H del aminoácido (n) puede establecer un enlace de hidrógeno con el grupo C=O del aminoácido (n+4). De esta forma, cada aminoácido (n) de la hélice forma dos puentes de hidrógeno con su enlace peptídico y el enlace peptídico del aminoácido en (n+4) y en (n-4). En total son 7 enlaces de hidrógeno por vuelta. Esto estabiliza enormemente la hélice.Esta dentro de los niveles de organización de la proteína.[31]
En 1868 Pierre Janssen y Joseph Norman Lockyer al analizar el espectro de la luz solar durante un eclipse solar ocurrido aquel año, encontraron una línea de emisión de un elemento desconocido. Eduard Frankland confirmó los resultados de Janssen y propuso el nombre helium para el nuevo elemento, en honor al dios griego del sol (Helios) al que se añadió el sufijo -ium ya que se esperaba que el nuevo elemento fuera metálico.
En 1895 Sir William Ramsay aisló el helio descubriendo que no era metálico, a pesar de lo cual el nombre original se conservó. Los químicos suecos Nils Langlet y Per Theodor Cleve consiguieron también, por la misma época, aislar el elemento.
En 1907 Ernest Rutherford y Thomas Royds mostraron que las partículas alfa son núcleos de helio. En 1908 el físico alemán Heike Kamerlingh Onnes produjo helio líquido enfriando el gas hasta 0,9 K, lo que le hizo merecedor del premio Nobel. En 1926 su discípulo Willem Hendrik Keesom logró por vez primera solidificar el helio.
El helio es el segundo elemento más abundante del universo tras el hidrógeno y constituye alrededor del 20% de la materia de las estrellas, en cuyo proceso de fusión nuclear desempeña un importante papel. La abundancia de helio no puede ser explicada por la generada en las estrellas, aunque es consistente con el modelo del Big bang, creyéndose que la mayor parte del helio existente se formó en los tres primeros minutos del universo.
En la atmósfera terrestre hay del orden de 5 ppm y se encuentra también como producto de desintegración en diversos minerales radioactivos de uranio y torio. Además está presente en algunas aguas minerales, en gases volcánicos y en ciertos yacimientos de gas natural de los Estados Unidos, de los que proviene la mayoría del helio comercial.
El helio puede sintetizarse bombardeando núcleos de litio o boro con protones a alta velocidad.
Dado que el helio es un gas noble, en la práctica no participa en las reacciones químicas, aunque bajo la influencia de descargas eléctricas o bombardeado con electrones forma compuestos con el wolframio, yodo, flúor y fósforo.
Isótopos
Representación esquemática de un átomo de 4He.El isótopo más común del helio es el 4He, cuyo núcleo está constituido por dos protones y dos neutrones. Su excepcional estabilidad nuclear se debe a que tiene un número másico de nucleones, es decir, una cantidad que se distribuye en niveles completos (de modo análogo a como se distribuyen los electrones en los orbitales). Numerosos núcleos pesados se desintegran emitiendo un núcleo de 4He; éste proceso, que se denomina desintegración alfa y por el que al núcleo emitido se le llama partícula alfa, es el origen de la mayoría del helio terrestre.
El helio tiene un segundo isótopo, el 3He, así como otros más pesados que son radiactivos. El helio-3 es prácticamente inexistente en la tierra, dado que la desintegración alfa produce exclusivamente núcleos de helio-4 y tanto éstos como el helio atmosférico escapan al espacio en periodos geológicos relativamente cortos.
Ambos isótopos se produjeron en el Big bang y cantidades significativas se siguen produciendo mediante la fusión del hidrógeno en las estrellas siguiendo la cadena protón-protón.[32]
G0RD0N escribió:Respecto al párrafo resaltado, es sin embargo donde más se le va la castaña y en definitiva tampoco veo dónde quiere llegar. Primero habla de que en la síntesis del átomo se libera energía, con lo cual habla de la fusión nuclear, pero luego se sirve de este hecho para dilucidar sobre la descomposición atómica del cuerpo, que en realidad sería fisión nuclear; no sé, no le veo mucho sentido o quizá es que ya estoy espeso . Y lo de que no poder crear neutrones en el átomo a partir de radiación, pues yo tampoco conozco ninguna. Sin embargo, lo que sí que usamos es la creación de neutrones en según qué átomos a partir de otros neutrones. Por ejemplo, en fusión usamos Berilio como multiplicador de neutrones y lo bombardeamos con un neutrón (tanto da que sea lento como rápido ya que su "susceptibilidad" para absorver neutrones -técnicamente cross-section- es alta) para obtener 2, aunque esto conlleva la transmutación del Berilio a Helio. A pesar de ello, este Helio es inestable con estos 2 neutrones, con lo que son expulsados del átomo para dar el Helio estable que todos conocemos.
La cadena protón-protón es una de las dos reacciones de fusión que se producen en las estrellas para convertir el hidrógeno en helio, el otro proceso conocido es el ciclo CNO. Las cadenas protón-protón son más importantes en estrellas del tamaño del Sol o menores. El balance global del proceso es el equivalente de unir cuatro nucleones y dos electrones para formar un núcleo de helio-4 (2 protones + 2 neutrones).[33][34]
El proceso triple alfa (TPA) es el proceso por el cual tres núcleos de helio (partículas alfa) se transforman en un núcleo de carbono.
Esta reacción nuclear de fusión sólo ocurre a velocidades apreciables a temperaturas por encima de 100 000 000 kelvin y en núcleos estelares con una gran abundancia de helio. Por tanto, éste proceso sólo es posible en las estrellas más viejas, donde el helio producido por las cadenas protón-protón y el ciclo CNO se ha acumulado en el núcleo. Cuando todo el hidrógeno presente se ha consumido, el núcleo se colapsa hasta que se alcanzan las temperaturas necesarias para iniciar la fusión de helio.
4He + 4He ↔ 8Be
8Be + 4He ↔ 12C + γ + 7.367 MeV
El 8Be producido durante la primera etapa es muy inestable y decae otra vez en dos núcleos de helio en 2.6·10-16 segundos. De todas formas, en las condiciones en las que se fusiona el helio siempre hay pequeñas cantidades de 8Be presentes en equilibrio; la captura de otra partícula alfa da lugar al 12C. El proceso global de conversión de tres partículas alfa en un núcleo de 12C se denomina proceso triple alfa.
Ya que dicho proceso es improbable, debido a la escasa cantidad de 8Be presente en un momento dado, se necesita muchísimo tiempo para formar carbono. Como consecuencia no se produjo carbono durante el Big Bang, ya que la temperatura descendió a niveles inferiores a los requeridos para que se dé ésta reacción.
La siguiente etapa donde el oxígeno formado se combina con otra partícula alfa para dar lugar a neon es más dificultosa, debido a las reglas de espín nuclear, y por tanto no pueden formarse elementos más pesados por esta vía.
Como resultado de estas reacciones, se forman grandes cantidades de carbono y oxígeno pero sólo fracciones diminutas de éstos se transforman en neón y otros núcleos más pesados, siendo por tanto éstos dos las principales cenizas de la combustión del helio. Las resonancias nucleares que dan lugar a tales cantidades de carbono y oxígeno se citan generalmente como evidencia del principio antrópico.
Las reacciones de nucleosíntesis por fusión nuclear sólo producen elementos hasta el 56Fe, el núcleo atómico más estable; los elementos más pesados se producen por procesos captura neutrónica. La captura lenta, el proceso S, produce aproximadamente la mitad de dichos elementos. La otra mitad se produce en el proceso R o captura rápida, proceso que probablemente tenga lugar en el núcleo de las supernovas de colapso.
El proceso triple alfa (TPA) depende fuertemente de la temperatura y la densidad del material estelar. La energía liberada en ésta reacción es aproximadamente proporcional a T30 y al cuadrado de la densidad. En comparación, las cadenas protón-protón (PP) liberan energía proporcionalmente a la cuarta potencia de la temperatura y en proporción directa a la densidad.
Esta fuerte dependencia de la temperatura tiene consecuencias en la última etapa de la evolución estelar, la de gigante roja.
Para masas estelares menores, el helio acumulado en el núcleo previene el colapso estelar mediante la presión de degeneración electrónica (véase Principio de exclusión de Pauli). Por tanto, el volumen del núcleo depende sólo de la densidad y no de la presión. Una consecuencia de este hecho es que una vez que una estrella pequeña ha llegado a éste estado, sólo puede ir aumentando la temperatura del núcleo hasta que se llega al punto de ignición del helio. Ya que la velocidad de este proceso depende fuertemente de la temperatura, y sin una expansión estelar que la disminuya, dicha velocidad aumenta exponencialmente, consumiéndose del 60 al 80% del helio presente en pocos minutos. En ese momento se liberan cantidades inmensas de energía cuando la alta temperatura provoca la expansión repentina de la estrella, dando lugar al llamado "flash de helio" [35]
El gluón (de la voz inglesa glue 'pegamento', derivada a su vez del latín glūten a través del francés gluer 'pegar') es el bosón portador de la interacción nuclear fuerte, una de las cuatro fuerzas fundamentales. No posee masa ni carga eléctrica, pero sí carga de color, por lo que además de transmitir la interacción fuerte también la sufre.
Al igual que el fotón, el gluón es un bosón sin masa ni carga de spin 1. Como los quarks, los gluones tienen carga de color, que depende del cambio de color de los quarks. Los quarks cambian de color cuando se intercambian gluones, de tal forma que la carga de color total del sistema formado por el quark y el gluón, antes y después de la emisión o absorción es la misma.
La teoría que postula la existencia de los gluones y describe su dinámica se llama cromodinámica cuántica. El nombre hace alusión a "pegamento" (glue), se debe a estas partículas son las que "unen" los quarks dentro de los nucleones.
Al sufrir ellos mismos su propia interacción, los gluones que unen los quarks crean un campo de color con forma de cuerda que impide que los quarks se separen con una fuerza inmensa. La formación de éstas estructuras con forma de cuerda por parte de los gluones limita el campo de acción de ésta interacción a un orden de 10-15 metros (más o menos el tamaño de un núcleo atómico).
Al contrario que la fuerza eléctrica o la gravitatoria, si se intenta separar entre sí un par de quarks, el campo de color tira de ellos con mucha más fuerza; es como si los quarks estuvieran unidos por un "muelle gluónico", que intenta volver a su longitud inicial. Debido a esto, los quarks y los gluones son partículas muy difíciles de detectar y sólo podemos ver las partículas que ellos forman, los hadrones.
Cuando se separan tanto dos quarks unidos mediante éste muelle, se acumula tanta energía en el sistema que es más fácil para el mismo crear nuevos quarks para devolver el campo de color a un estado menos energético. Esto es resultado de convertir parte de la energía del campo de color en nueva materia (E = mc2).[36][37]
Disco de acrecimiento
Los sistemas planetarios se originan a partir de discos de este tipo mediante fenómenos de acrecimiento (o agregación) de las partículas originarias, hasta formar los planetas, satélites y los cuerpos menores del sistema.
A modo de ejemplo, cuando se deja caer miel lentamente desde un envase, se crea una especie de disco de acrecimiento entre la Tierra y la miel del envase, debido a la gravedad. Esto es, básicamente, un disco de acrecimiento ya que se forma con principios similares en cuestiones gravitatorias. Podría decirse que, en la Tierra, la consistencia de la miel (y líquidos similares) posee un comportamiento parecido al de la masa estelar que constituye un disco de acrecimiento, el cual está hecho del plasma que compone las estrellas.
Una estrella u otro astro situado en un sistema binario puede también formar un disco de acrecimiento robando materia de las capas exteriores de su compañera. Esta materia forma un anillo en torno a la estrella captora, pudiendo llegar a caer sobre la superficie de la misma tras describir una trayectoria en espiral. Debido a las enormes velocidades que alcanza la materia en dicha caída se observa una fuerte emisión de rayos X, que ha servido para detectar objetos que no emiten radiación por sí solos, como es el caso de las estrellas de neutrones o los agujeros negros. Estos sistemas binarios se conocen como binarias de rayos X.
El disco es una estructura común en el universo. Tanto galaxias como estrellas se han formado a la vez en discos de acrecimiento de muy diferentes dimensiones. El motivo que origina tan comunes estructuras a partir de informes nubes de gas es sencillo. Casi toda masa de gas posee un cierto momento angular, una mínima cantidad de rotación. Es decir, las inmensas nubes que se colapsan formando estas estructuras giran inicialmente, aunque sea muy lentamente. El sistema de gas en rotación se mantiene en un delicado equilibrio que se puede romper debido, p.ej., a la onda de presión de una supernova o a que alcanza una cantidad de masa crítica. Cuando sobreviene la inestabilidad y la nube se comprime por el efecto creciente de la gravedad ésta empieza a experimentar ciertos cambios que la conducirán a formar un disco.
Al comprimirse la nube gira más deprisa por conservación del momento angular. Pero este giro solo ocurre a lo largo de su plano de giro. En las zonas de mayor rotación la fuerza centrífuga adquiere cada vez mayor intensidad. Esta asimetría cada vez más acusada es la que, poco a poco, acaba por dar forma al disco. Las regiones suprayacentes y subyacentes al plano de giro, es decir los polos, caen libres a gran velocidad mientras que el gas que gira a lo largo de dicho plano se ve muy frenado por la creciente fuerza centrífuga. Así pues, la acción combinada de rotación y gravedad es la que, al final, dará la característica forma de disco.
Los discos de acrecimiento más activos presentan fuertes chorros de emisión de material a lo largo del eje de rotación. Este fenómeno se denomina comúnmente difusión ambipolar. La estructura y naturaleza de los mecanismos de emisión en chorro no se conocen con precisión aunque se cree que tienen que ver con la presencia de un fuerte campo magnético. El material central fuertemente ionizado escupe una parte de sí a través de las líneas de campo que actúan a modo de guías.
A menudo, en sistemas binarios en los que una de las estrellas es un objeto compacto como un púlsar o un agujero negro las observaciones muestran indicios de material circulando desde la estrella brillante hacia el objeto compacto. Esto ocurre cuando la estrella posee sus capas exteriores en el interior del límite de Roche del objeto compacto. El material arrancado fluye sobre dicho objeto formando un disco de acrecimiento a su alrededor. En el caso de los agujeros negros, la materia se llega a acelerar tanto que las emisiones de radiación procedentes del vórtice se dan en la banda de los rayos X. Las fuentes de rayos X suelen ser, de hecho, una pista que delata su presencia.
http://es.wikipedia.org/wiki/Disco_de_acrecimiento
La resonancia magnética nuclear (RMN)es un fenómeno físico basado en las propiedades mecánico-cuánticas de los núcleos atómicos. RMN también se refiere a la familia de métodos científicos que explotan este fenómeno para estudiar moléculas (espectroscopia de RMN), macromoléculas (RMN biomolecular), así como tejidos y organismos completos (imagen por resonancia magnética).
Podría parecer por lo dicho arriba que todos los núcleos del mismo núclido (y por lo tanto la misma γ) resuenan a la misma frecuencia. Este no es el caso. La perturbación más importante en las frecuencias de RMN para aplicaciones en RMN es el efecto de 'apantallamiento' que ejercen los electrones circundantes. En general, este apantallamiento electrónico reduce el campo magnético del núcleo (lo cual determina la frecuencia de la RMN). Como resultado, la brecha energética se reduce y la frecuencia requerida para alcanzar resonancia también se reduce. Este desplazamiento de la frecuencia de RMN dado por el ambiente químico se conoce como desplazamiento químico, y explica porque el RMN es una sonda directa de la estructura química. Si un núcleo esta mas apantallado, estará desplazado hacia 'campo alto' (menor desplazamiento químico) y si esta mas desapantallado, entonces estará desplazado hacia 'campo bajo' (mayor desplazamiento químico).
A menos que la simetría local sea particularmente alta, el efecto de apantallamiento depende de la orientación de la molécula con respecto al campo externo. En RMN de estado sólido, el 'giro al ángulo mágico' (magic angle spinning) es necesario para disipar esta dependencia orientacional. [38]
Knos escribió:Vale... Que tiene que ver esto con la sabana santa entonces?
Moki_X escribió:Killbastards escribió:Bajo estas y otras premisas podemos a llegar a comprender que en el universo existen 2 tipos de electromagnetismo, el electromagnetimo centrípato y el centrífugo. El electromagnetismo centrípato tiene que ver con las correlaciones cuanticas propias y el electromagnetismo centrífugo tiene que ver con las correlaciones cuánticas generales.
Joder... hasta aqui he llegado a leer.
Bajo esta y otras premisas... ¿Cuales son esas otras premisas?
Define electromagnetismo centrípato y centrífugo. Porque no lo has definido ni has dicho en qué consiste.
Ansioso espero a que me respondas.
, no había visto semejante cosa." KillBastardsII escribió:Tocho que te cagas, que no incluye la definición de electromagnetismo centrípato y centrífugo y que empieza hablando de la visión y termina barruntando cosas acerca de la sábana santa
Moki_X escribió:Tio... tu eres un troll. Si o si
Moki_X escribió:Ahora mismo no estás manteniendo un diálogo en un foro de discusión, sino que estás soltando verborrea sin atender a lo que te dicen los demás.
KillBastardsII escribió:Creo que ya te respondí en el principio del todo del hilo "tocho" de más arriba. O es que ni siquiera has leído los principios de mis mensajes...
KillBastardsII escribió:. Antes me habéis dicho que digo cosas inconexas pero no me habéis sabido concretar donde están las inconexiones o la información que no tiene nada que ver con el tema general del hilo.
KillBastardsII escribió:Lo que me refiero es a que solo hay 2 tipos de fuerza o electromagnetismo que conozcamos actualmente;
Energía centrípeta: Energía que busca el centro.
Energía centrifuga: Energía que huye del centro.
KillBastardsII escribió:Bajo esta y otras premisas... ¿Cuales son esas otras premisas?
Define electromagnetismo centrípato y centrífugo. Porque no lo has definido ni has dicho en qué consiste.
Lo que me refiero es a que solo hay 2 tipos de fuerza o electromagnetismo que conozcamos actualmente;
Energía centrípeta: Energía que busca el centro.
Energía centrifuga: Energía que huye del centro.
A continuació expongo la segunda parte del mensaje:Kyubi chan escribió:killbastards tio, que yo no hago la fotosíntesis, y mi perro tampoco. No le des más vueltas.
Di que te has equivocado y no pasa nada, se puede seguir debatiendo.
Yo no me equivoqué en decir que los humanos hacemos la fotosíntesís con la luz del sol. Si me equivocado en algo, es en no haber sido lo suficientemente dinámico y rápido como para no producir malos entendidos o malas interpetraciones sobre lo que expuse anteriormente. Por eso mismo, voy a tratar de explicar bien en que me fundamento.
Antes dije que los humanos hacemos fotosintesis de diferentes maneras. Una de ellas mediante la vision. En este punto me gustaría detallar un poco como es este proceso:Cita de la wikipedia sobre la vision
La visión ocular es uno de los procesos fotosintéticos más complejos que conocemos del universo...Y el proceso de fotosíntesis que se produce en la visión ocular, de alguna forma, esta correlacionado con el efecto fotosintético de la imagen de la sábana santa y el efecto fotosintético general.
Cuando hubo el primer pecado de nuestros padres Adam y Eva, ellos y nosotros fuímos condenados a no residir desde el principio del todo en el paraiso. De forma alguna, el efecto de salida de nuestros antepasados del paraiso para venir a existir en la tierra por culpa del pecado, esta correlacionado con el efecto que se produjo en la sábana santa. Jesucristo redime con su resurección, el primer pecado que inhabilita a los humanos poder existir en el cielo. El efecto de la imagen de la sábana santa esta correlacionado con la salida del paraiso de nuestros padres e inicio de vida en la tierra y este efecto de la imágen de la sábana santa también esta correlacionado con el juicio final de todos los tiempos. La curvisa es de Moki_X (o séase, mia): Bueno, bueno, es que esto ya no tiene nada que ver ni con el electromagnetismo, ni con la física de partículas, ni con la termodinámica, ni con nada. Sentido 0
Ahora también, para seguir buscando correlaciones cuánticas con la sábana santa y lo explicado anteriormente voy a colgar más información sobre el origen de los astros y el espacio:Cita de la wikipedia sobre astrofísicaG0RD0N escribió:Respecto al párrafo resaltado, es sin embargo donde más se le va la castaña y en definitiva tampoco veo dónde quiere llegar. Primero habla de que en la síntesis del átomo se libera energía, con lo cual habla de la fusión nuclear, pero luego se sirve de este hecho para dilucidar sobre la descomposición atómica del cuerpo, que en realidad sería fisión nuclear; no sé, no le veo mucho sentido o quizá es que ya estoy espeso . Y lo de que no poder crear neutrones en el átomo a partir de radiación, pues yo tampoco conozco ninguna. Sin embargo, lo que sí que usamos es la creación de neutrones en según qué átomos a partir de otros neutrones. Por ejemplo, en fusión usamos Berilio como multiplicador de neutrones y lo bombardeamos con un neutrón (tanto da que sea lento como rápido ya que su "susceptibilidad" para absorver neutrones -técnicamente cross-section- es alta) para obtener 2, aunque esto conlleva la transmutación del Berilio a Helio. A pesar de ello, este Helio es inestable con estos 2 neutrones, con lo que son expulsados del átomo para dar el Helio estable que todos conocemos.
Gordon, yo tampoco entiendo muy bien el proceso exacto de descomposición para que se de una imagen como la de la sábana santa, ni tampoco entiendo del todo bien como funciona en todas sus variantes el proceso de radiación. Pero veo correlaciones entre el proceso de la imagen con el proceso y fin de la creación de la vida, sin llegar a entender bien todos los entresijos de estos temas.
Pero sin embargo dilucido que en el caso de conocer bien el proceso y los motivos de la creación y el fin de la vida en la tierra, se puede entender el proceso de la creación de la imagen de la sábana santa. Repito que yo no las conozco bien del todo, pero si veo correlaciones cuánticas sobre ambos procesos.
Vayamos pues a poner más información sobre estas correlaciones:Cita de la wikipedia sobre Física nuclear y de partículasKnos escribió:Vale... Que tiene que ver esto con la sabana santa entonces?
La imagen de la sábana santa es un tipo de imagen como de "resonancia magnética universal" imprimida sobre una superficie espacial relativa. Utilizando las técnicas más prodigiosas para reestructurar un cuerpo a partir de principios antropológicos universales.
"Sé de un hombre en Cristo, el cual hace catorce años -si en el cuerpo o fuera del cuerpo no lo sé, Dios lo sabe- fue arrebatado hasta el tercer cielo. Y sé que este hombre -en el cuerpo o fuera del cuerpo del cuerpo no lo sé, Dios lo sabe- fue arrebatado al paraíso y oyó palabras inefables que el hombre no puede pronunciar” (2Co 12,2-4)
Dijo San Pablo, hombre que conoció a Cristo en persona y conoció su resurrección y de alguna manera, conocía todo lo que implicaba la resurrección de Jesucristo.
"El orden de la razón práctica es correlativo al orden de la razón correlativa".
Dijo Santo Tomas de Aquino hace muchos siglos atrás. Ambos dichos de estos santos de la iglesia, son afirmaciones que se correlacinan entre sí, entre lo información que intento correlacionar con este mensaje, entre la fotosíntesis y entre la imagen de la sábana santa.

Skalextric escribió:Partiendo de que la fuerza centrífuga no es una fuerza real como tal, mezclas muchas tonterías. ¿Electromagnetismo centrífugo? Por favor...
Y sí, edita tu post porque tío ahí no decía nada de esa fuerza que nombras.
El universo nace en circunstancias desconocidas. Según los conocimientos científicos del Bing Bang, surgió de una “singularidad”, un punto de densidad infinita en el que explotan las leyes del espacio y del tiempo.
http://html.rincondelvago.com/historia- ... verso.html
Durante su formación, un estrella experimenta diferentes fases antes de alcanzar la estabilidad que le proporciona la generación de energía a partir de la combustión de hidrógeno en su interior (lo que se denomina Secuencia Principal). En primer lugar, una nube interestelar de polvo y gas se fragmenta y colapsa debido a instabilidades, dando lugar a la aparición de varios nucleos. Más tarde, las protoestrellas aparecen a partir de estos nucleos que continuan con su colapso. Algo más tarde, la conservación del momento angular hace que se forme un disco alrededor del objeto central, que proporciona el material a la estrella (mediante el acrecimiento). Finalmente, el disco termina por desaparecer, pero un sistema protoplanetari se puede formar durante esta última etapa. La fase de acrecimiento corresponde a los objetos de tipo TTauri clásica o Clase II. Esto objetos muetran una peculiaridades observacionales características, como puede ser excesos en el ultravioleta o el infrarrojo, emisiones intensas en determinadas líneas como Halfa. Suele durar unos pocos millones de años, una fracción muy reducida de la vida total de la estrella, pero de vital imprtancia.
En la actualidad nos e conoce con exactitud el mecanismo de formación de las enanas marrones, pero las evidencias observacionales indican que se forman de manera análoga a las estrellas, por colapso y fragmentación de la nube de material interestelar de polvo y gas. Diferentes grupos, entre los que nos encontramos nosotros, han encontrado excesos infrarrojos en objetos subestelares (especialmente con el satélite de NASA Spitzer), intensísimas emisiones en H(alfa), en algunos casos con líneas de gran anchura (lo que indica claramente queson debidas al acrecimiento). También se han detectado líneas prohibidas producidas por los chorros de materia expulsados desde la parte más interna del disco, tales como OIII6300&6364, NII 6548&6581, SII 6717&6731.
http://www.laeff.inta.es/index.php?op=m ... recimiento
El universo surgió de una "singularidad", un punto de densidad infinita(fuerza centrípeta) en el que explotan las leyes del espacio y del tiempo(fuerza centrífuga). O también se le puede llamar efecto de colapso y fragmentación. Antes de que surgiera la gravedad y las energías nucleares que conocemos, antes de todo eso se produjo el efecto de colapso y fragmentación.
KillBastardsII escribió:Bueno pues llamalo efecto de colapso y fragmentación, haber si así se entiende mejor. Y así no te suena a tontería. Yo tampoco uso las mejores expresiones lo reconozco.
KillBastardsII escribió:Hay 4 fuerzas elementales: gravedad, electromagnetismo, fuerza nuclear debil y fuerza nuclear fuerte.
De lo poco que sabemos sobre el origen de el universo es que, las 2 fuerzas primogénitas antes de que se diera el Bigbang y la rotura del primer disco de acrecimiento universal, son la fuerza centrípeta y la fuerza centrífuga¡Es que eso no es contestar! Te pido que me definas Electromagnetismo centrípeto y me dices "es una energía que busca el centro". Eso no es una definición de nada. Sólo es una afirmación categórica sin ningún tipo de referencias. ¿Eres capaz de citarme algún libro de Física en el que se defina el electromagnetismo centrípeto/centrífugo?. Me juego mi NES a que no.
Supongo que te servirán estos enlaces sobre información astrofísica:Diagrama de una estrella o enana marrón con disco y accrecimiento.
Durante su formación, un estrella experimenta diferentes fases antes de alcanzar la estabilidad que le proporciona la generación de energía a partir de la combustión de hidrógeno en su interior (lo que se denomina Secuencia Principal).En primer lugar, una nube interestelar de polvo y gas se fragmenta y colapsa debido a instabilidades, dando lugar a la aparición de varios nucleos. Más tarde, las protoestrellas aparecen a partir de estos nucleos que continuan con su colapso. Algo más tarde, la conservación del momento angular hace que se forme un disco alrededor del objeto central, que proporciona el material a la estrella (mediante el acrecimiento). Finalmente, el disco termina por desaparecer, pero un sistema protoplanetari se puede formar durante esta última etapa. La fase de acrecimiento corresponde a los objetos de tipo TTauri clásica o Clase II. Esto objetos muetran una peculiaridades observacionales características, como puede ser excesos en el ultravioleta o el infrarrojo, emisiones intensas en determinadas líneas como Halfa. Suele durar unos pocos millones de años, una fracción muy reducida de la vida total de la estrella, pero de vital imprtancia.
En la actualidad nos e conoce con exactitud el mecanismo de formación de las enanas marrones, pero las evidencias observacionales indican que se forman de manera análoga a las estrellas, por colapso y fragmentación de la nube de material interestelar de polvo y gas. Diferentes grupos, entre los que nos encontramos nosotros, han encontrado excesos infrarrojos en objetos subestelares (especialmente con el satélite de NASA Spitzer), intensísimas emisiones en H(alfa), en algunos casos con líneas de gran anchura (lo que indica claramente queson debidas al acrecimiento). También se han detectado líneas prohibidas producidas por los chorros de materia expulsados desde la parte más interna del disco, tales como OIII6300&6364, NII 6548&6581, SII 6717&6731.
http://www.laeff.inta.es/index.php?op=modload&name=phpWiki&file=index&pagename=Research-Brown%20Dwarfs-BD_esp-DiscosAcrecimiento
También te dejo un enlace a un libro de la libreria de google que habla sobre la formación de los discos de acreción que es el horizonte donde la materia energía y fuerza puede tomar solo dos direcciones o hacia denntro del disco o hacia fuera del disco, ese momento donde la energía gira en el disco de acreción es el preludio de escoger una de las dos direcciones posibles o hacia el centro o hacia fuera del disco de acreción. El disco de acreción esta correlacionado con el principio de la existencia del universo.
http://books.google.es/books?id=muM-V-gJbUgC&pg=PA409&dq=disco+acrecion&cd=6#
Tu mismo has dicho que mi explicación era categórica y sin referencias, bueno, las referencias las puse en el anterior post cuando hablé sobre los discos de acreción. Ahora ya te he detallado un poco más el funcionamiento de estas 2 tipos de fuerzas primordiales.
PD: Me parece que me ganado tu Nes. A no ser que sepas tumbar estos argumentos astrofísicos.
Saludos.
zheo escribió:Killbastards, ¿fuiste tu al que contrataron para escribir el contenido de la web de PowerBalance?

death77 escribió:Joder cómo la liáis.
Si Dios existe, es un cabrón rencoroso egoísta y vago por dejar que el mundo cada vez esté tan mal.
Y si no existe, este hilo es inútil
No sé qué es peor
Moki_X escribió:Define tu fuerza electromagnética centrípeta
KillbastardsII escribió:"oh, vamos a hablar de estrellas y discos de acreción, a ver si cuela"
Moki_X escribió:¿A dónde vas?
KillbastardsII escribió:Manzanas traigo
Skalextric escribió:Y lo bonito que es sacar a la gente de la ignorancia... lo digo sin maldad, pero si alguien tiene conocimientos y los pone al servicio de otros para que vean los errores que comenten no está tan mal, ahora si ese otro no quiere bajarse del burro no hay mas remedio. También digo que algunos vamos a veces de prepotentes y mas de un post he leido así (incluido alguno mío, aunque lo evito en la medida de lo posible).
![[boing]](/images/smilies/nuevos/pelota_ani1.gif "pelota")
![[buuuaaaa]](/images/smilies/nuevos/triste_ani3.gif "a lágrima viva")
![[mamaaaaa]](/images/smilies/nuevos/triste_ani4.gif "llorica")
Elegido escribió:Supongo que no has podido aguantar la vergüenza de soltar tal cantidad de tonterías que ni tú mismo te las creías. Por caridad, te aclaro unas cuestiones :
Elelegido escribió:1. La ciencia no sabe nada acerca de cualquier periodo temporal anterior al Big Bang, ni siquiera de los primeros instantes tras su surgimiento. Tanto es así, que tampoco se puede afirmar que el "tiempo anterior al Big Bang" sea parte de la naturaleza del Cosmos.
El astrónomo norteamericano Edwin Hubble descubrió en 1938 que todas las galaxias se alejan entre sí a gran velocidad. Ellos sirvió de base para que el físico George Gamow propusiera en 1949 la teoría del Big Bang (o la Gran Explosión) para explicar el origen del universo.
Según Gamow, todo lo que vemos se formó a partir de una concentración muy densa (prácticamente un punto geométrico) de materia que “estalló” y desde entonces ha ido enfriándose y aumentando de volumen, mientras se fueron formando las partículas, los átomos, las estrellas, los planetas…
http://jaespimon.wordpress.com/tag/fuerza-centripeta/
2. Las únicas 4 fuerzas que se conocen, son el electromagnetismo, la nuclear débil, la nuclear fuerte y la gravedad. Una fuerza es una interacción que cambia la cantidad de movimiento de una partícula. Y por tanto, la fuerza centrípeta o centrífuga en un contexto dado es resultado de una aplicación específica de esas cuatro fuerzas. Además, la tendencia lleva a investigar si esas cuatro fuerzas son diferentes caras de una única fuerza a base de unificaciones. Ya se ha realizado un modelo con razonable éxito unificando la fuerza electromagnética y la nuclear débil conformando la fuerza electrodébil.
3. La energía no puede tener "direcciones". Es un escalar, y una propiedad de la materia y del espacio.
4.Cuando hablas de direcciones, quieres decir sentido.
5. Para cualquier estructura geométrica que represente un ente en el Universo, se puede aplicar la misma dicotomía de "O va hacia dentro, o va hacia fuera". Es más, afirmo que en el universo todas las partículas o van hacia mi pene, o no van hacia mi pene, sin miedo a equivocarme.
6. La energía no gira.
7. La energía no escoge.
8.Un disco de acreción no es más que un cúmulo de materia bajo la influencia de un campo gravitatorio comparativamente muy grande. La materia arrastrada tiende a ir al cuerpo que provoca dicho campo, del mismo modo que actúa la fuerza gravitatoria en todos los ámbitos de nuestra realidad cotidiana. Eso origina la estructura toroidal típica de un disco de acreción.
9.Todo lo que existe en el Universo está correlacionado con la existencia del Universo.
10. Has detallado varias cosas :
a) Que no puedes seguir adelante con esta argumentación.
b) Que eres bastante desconocedor de la ciencia.
c) Que atentas contra ella inventándote hipótesis sin ningún rigor científico y vendiéndolas como si lo tuvieran.
Pero no has detallado el funcionamiento de ningunas 2 fuerzas primordiales.
Killbastards, ¿me puedes decir dónde está la web que te saca fragmentos aleatorios de páginas de la wikipedia? Si, esa que estás usando
6.El movimiento circular uniforme es un caso “especial”, pues posee aceleración. Esto parece un contrasentido, ya que te preguntarás: ¿Cómo un movimiento uniforme puede tener aceleración?
Hay aceleración debido al cambio continuo de dirección del vector velocidad a lo largo de todo el movimiento.
Dicha aceleración está siempre dirigida hacia el centro, por lo que se llama aceleración centrípeta. Por otro lado, este vector puede verse que es perpendicular (o normal) al vector velocidad en todo momento. Por ello también se le denomina aceleración normal. Su módulo se obtiene dividiendo el cuadrado de la velocidad entre el radio de la trayectoria.
2. Te lo voy a explicar de otra manera, de hecho las 4 fuerzas que comentas se pueden divivir entre 2 grupos de referencia.
Efecto centrípeto: Fuerza nuclear debil y fuerte; Unión con el medio universal
Efecto centrífugo: Efecto electromagnético y fuerza gravedad; Unión con el estado.
Vuelvo a pedir disculpas si anteriormente no me explique lo suficientemente bien. Lo que te estoy diciendo es que las 4 fuerzas se pueden dividir en 2 grupos de efecto concreto.
Choper escribió:Ahora mismo tengo a Dios en mi ano, ¿ queréis hacerle alguna pregunta antes de que se vaya ?

![[poraki]](/images/smilies/nuevos/dedos.gif "por aquí!")